
Aglyptodactylus chorus, Köhler, Jörn, Glaw, Frank, Pabijan, Maciej & Vences, Miguel, 2015
|
publication ID |
https://doi.org/10.11646/zootaxa.4006.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:136F727A-1628-449F-B762-254371D27360 |
|
DOI |
https://doi.org/10.5281/zenodo.5684812 |
|
persistent identifier |
https://treatment.plazi.org/id/145C609A-10CF-46F9-BA14-C5431EF52E00 |
|
taxon LSID |
lsid:zoobank.org:act:145C609A-10CF-46F9-BA14-C5431EF52E00 |
|
treatment provided by |
Plazi |
|
scientific name |
Aglyptodactylus chorus |
| status |
sp. nov. |
Aglyptodactylus chorus View in CoL sp. nov.
Remarks. This species has been referred to as Aglyptodactylus sp. aff. madagascariensis “East” by Glaw & Vences (2007) and Gehring et al. (2010), and Aglyptodactylus sp. 2 by Vieites et al. (2009) and Wollenberg et al. (2011), and as Aglyptodactylus sp. Ca2 by Perl et al. (2014).
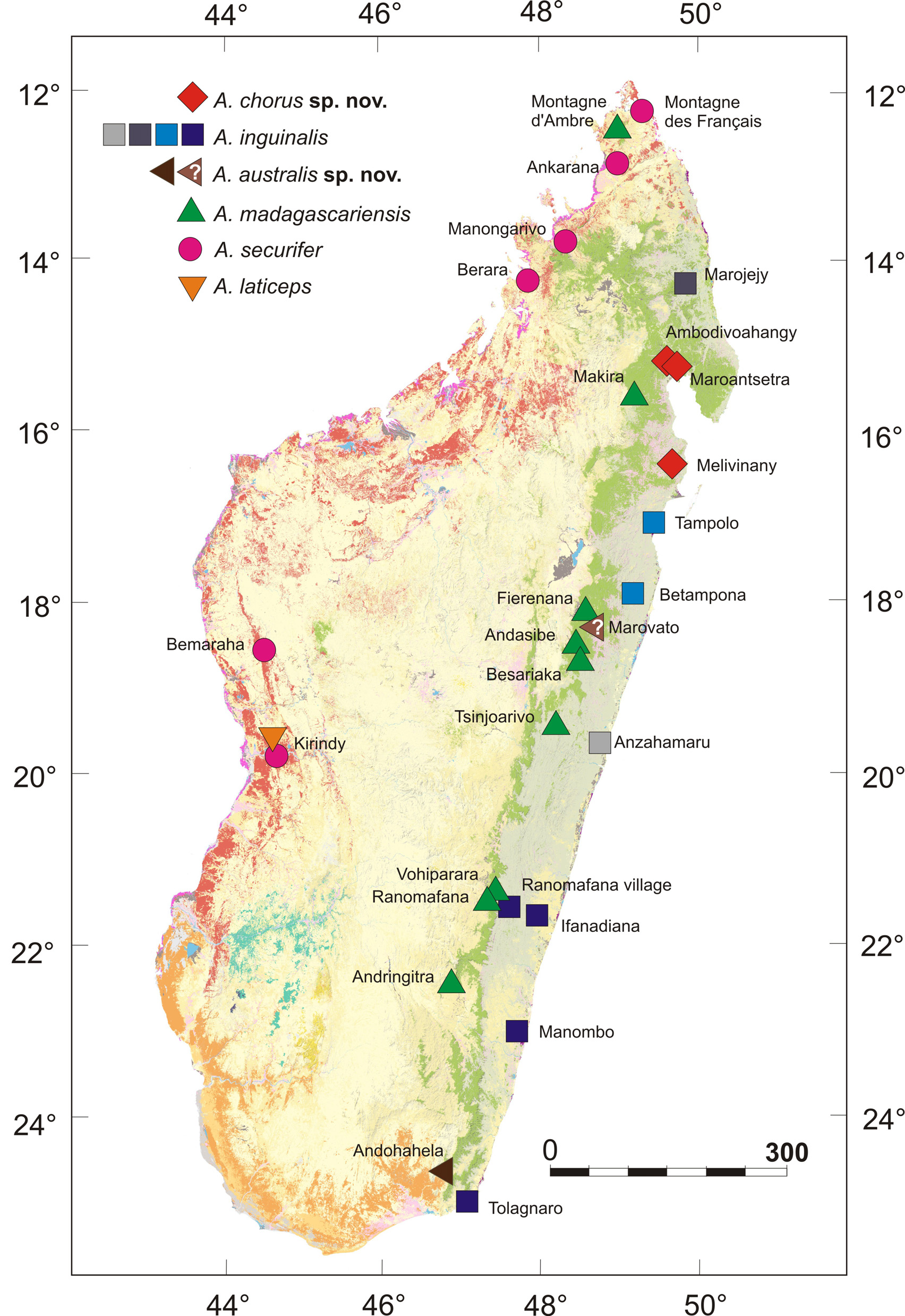
Holotype. ZSM 231/2010 (field number FGZC 4288), adult male, from Ambodivoahangy, 15°17'23.8'' S, 4937'13.0'' E, 24 m above sea level, Madagascar, collected on 3 April 2010 by P.-S. Gehring, F. Glaw, J. Köhler, M. Pabijan & F.M. Ratsoavina.
Paratypes. ZSM 230/2010 ( FGZC 4279), adult male, same data as holotype. ZSM 229/2010 ( FGZC 4233), adult female, collected on 31 March 2010, same locality and collectors as holotype. ZSM 140/2002 ( FGMV 2001.1415), adult male, ZSM 141/2002 ( FGMV 2001.1416), adult female, both from Maroantsetra, approximately 15°25’ S, 49°45’ E, 10 m above sea level, collected 16 December 2001 by M. Vences & A. Sarovy.
Referred specimens. ZSM 683/2009 ( ZCMV 11192), female, UADBA uncatalogued ( ZCMV 11193), juvenile, from Melivinany (a campsite in Befanjana forest; precise coordinates not available, but close to and west of the coastal village of Manompana, 16°41' S, 49°45' E), collected on 15 May 2009 by J.E. Randrianirina.
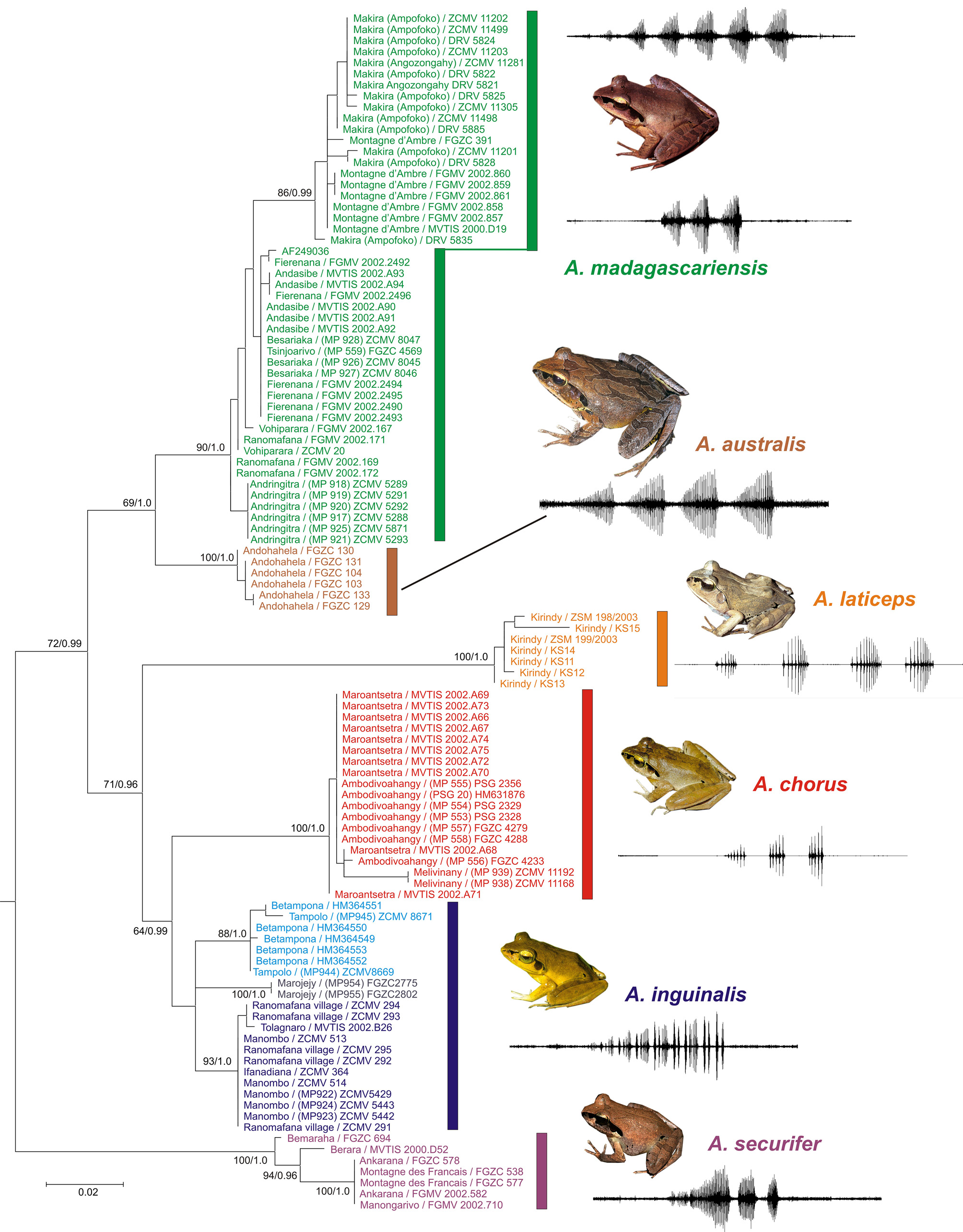
Diagnosis and comparison. A species of the genus Aglyptodactylus based on (a) digital terminal expansions not completely grooved, (b) black inguinal spots, (c) second finger shorter than first finger; (d) absence of femoral glands, (e) presence of nuptial pads in males, and (f) strong phenetic similarity to other Aglyptodactylus and molecular phylogenetic relationships.
Externally, the new species is characterized by the lack of a marbled pattern on posterior surfaces of thighs, lack of a vertebral line on dorsum, reduced dark lines and blotches in loreal region, well-developed webbing on feet, and a slender body. It differs from A. laticeps and A. securifer by a less broad head, more extensive webbing on feet, and a distinctly smaller inner metatarsal tubercle ( Fig. 4 View FIGURE 4 ). From A. madagascariensis , the new species differs by typically lacking a marbled pattern on posterior surfaces of thighs (present in most A. madagascariensis specimens), consistent absence of a fine vertebral line on dorsum (present in some specimens of A. madagascariensis ) and a slightly more pointed snout. According to the available data there is no external character reliably distinguishing A. chorus from A. inguinalis , but A. chorus differs from that species as from all other species in the genus by substantial genetic differentiation and advertisement call ( Figs. 1 View FIGURE 1 + 3; Tables 1 View TABLE 1 + 2).
Description of the holotype. Adult male ( Fig. 13 View FIGURE 13 a, b), as indicated by dark brown nuptial pads on the first finger. Intercalary element between ultimate and penultimate phalanges presumably present given the typical bentdown pattern of toe tips. Body slender; head slightly broader than body; snout pointed in dorsal view, nostrils directed dorsolaterally, distinctly nearer to snout tip than to eye; canthus rostralis straight and distinct, loreal region slightly concave, tympanum distinct, round, its horizontal diameter 67% of eye diameter; distinct supratympanic fold; tongue removed as tissue sample; vomerine teeth distinct in two well separated, round patches slightly behind the chonae, the diameter of the group of vomerine teeth about the same diameter as choanae. Arms slender, fingers with distinct single subarticular tubercles, outer metacarpal tubercle oval, flat; inner metacarpal tubercle only slightly elevated; fingers without webbing; comparative finger length 2<4<1<3, slightly developed terminal finger disks, without circummarginal grooves; limbs slender, tibiotarsal articulation reaches beyond snout tip if limb is adpressed along body; foot with an elongated inner metatarsal tubercle, no recognizable outer metatarsal tubercle, subarticular tubercle distinct, single, rounded; terminal toe disks slightly developed, lateral metatarsalia deeply separated, comparative toe length 1<2<3<5<4. Webbing formula 1(0.5), 2i (1), 2e (0), 3i (1.5), 3e (0), 4i (1.5), 4e (1), 5 (0). Skin on the upper surface smooth, throat, smooth, venter slightly granular. For morphological measurements see Table 3.
Colour of holotype in preservative (after 4 years in alcohol): head, dorsum, and dorsal surfaces of limbs greyish. An irregular interorbital dark marking, slightly extending onto the eyelids. Small brownish spots and elongated markings on the whole dorsum. Inguinal region with very small and inconspicuous black marking on each side. Flanks cream. Loreal and temporal region greyish (loreal region with small dark brown spots), tympanum brownish. A dark brown line on the supratympanic fold. An irregular dark brown blotch posterior to tympanum. Upper lip whitish. Dorsal surfaces of hands whitish, covered with minute grey spotting, dorsal surfaces of feet cream with fine greyish spotting; thighs and shanks with dark crossbands. Posterior surfaces of thighs with reticulated pattern. Cloacal region dark brown. Two or three narrow crossbands on feet. Ventral surfaces of feet grey. Arms greyish with one dark crossband on lower arm. A small black spot on the inner side of the lower arm. Posterior side of upper arm with some dark spotting. Iris blackish, pupil white. Belly and throat cream, anterior venter and ventral surfaces of arms cream.
In life, the general colour pattern was rather similar, but dorsal surfaces were yellowish cream with the impression of some golden tint. This yellow-golden tint was missing in the loreal and tympanal regions ( Fig. 13 View FIGURE 13 a). Venter was yellowish white, with the throat appearing somewhat darker. Ventral surfaces of thighs buff coloured, palmar and plantar surfaces buff with some dark brown spotting, nuptial pads dark brown ( Fig. 13 View FIGURE 13 b). Iris golden with some fine brown reticulation, inner iris area slightly darker. Iris periphery yellow, pupil black ( Fig. 13 View FIGURE 13 a).
Variation. For measurements of type specimens see Table 3. Male paratypes differ only slightly from the holotype in colouration. ZSM 230/2010 from the type locality has fewer dark dorsal markings compared to the holotype, larger dark spots in the loreal region, and lacks a dark interorbital marking. The topotypic female ZSM 229/2010 ( Fig. 13 View FIGURE 13 d) differs from the holotype by numerous distinct black spots on flanks, loreal, and tympanal region, lower lip cream with dark spotting and some greyish spots on venter. ZSM 140/2002 is brown on all dorsal surfaces with flanks being darker than the dorsum. The loreal region shows a dark brown band from eye to tip of snout. The anterior flanks are covered with a group of dark brown spots. ZSM 141/2002 is similarly brown dorsally and lacks the dark interorbital marking, but has a weakly defined dark Y-shaped marking on dorsum. The specimens from Melivinany (ZSM 682‒683/2009) have distinctly larger brown blotches on loreal and tympanal region. The male (ZSM 682/2009) has flanks and tympanal region covered with black spots, blackish crossbands on thighs and shanks, some greyish flecking on ventral surfaces of thighs and an indistinct reticulated pattern on posterior surfaces of thighs. The female (ZSM 683/2009) has a reddish brown dorsum, lacking distinct dark markings except for two dark flecks in the interorbital region.
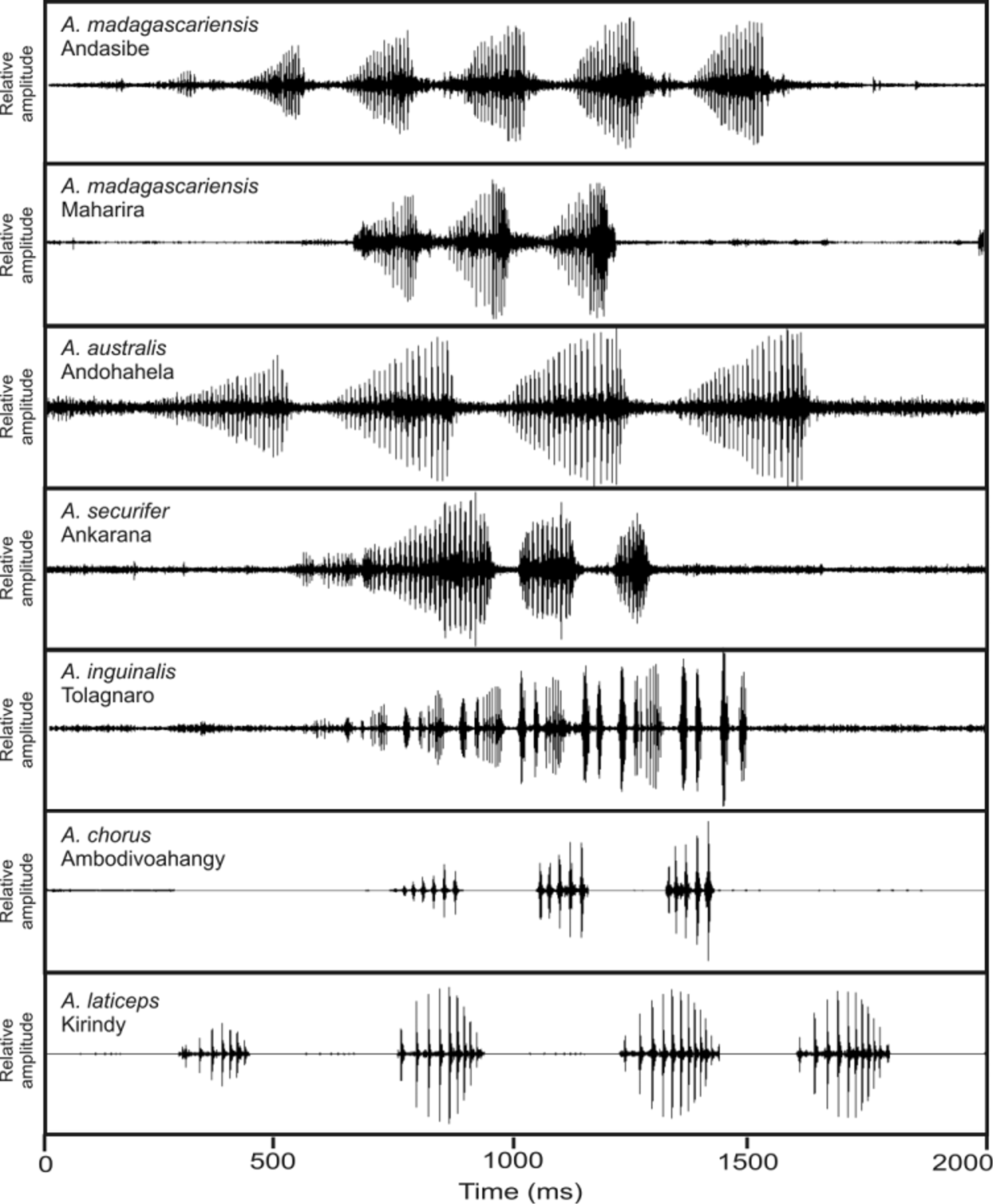
Call. Advertisement calls recorded at the type locality ( Fig. 14 View FIGURE 14 ) and almost certainty referable to the holotype ZSM 231/2010 were emitted in the context of a large chorus of males and were comparably soft. They consisted of 3‒4 distinctly pulsed notes with 73‒140 ms duration repeated at regular intervals. Each note is composed of 4‒6 pulses which are well-separated from each other and are repeated at a mean rate of 37.3 pulses per second. Amplitude increased from the first to the last note of the call. Within notes, amplitude slightly increased from first to last pulse of the note. Main call energy was distributed at approximately 900‒4000 Hz, with maximum call energy between 2756‒2820 Hz.
The call of A. chorus distinctly differs from the calls of A. australis , A. madagascariensis , A. securifer and A. inguinalis by lower pulse repetition rate. It furthermore differs from the first three mentioned species by shorter note duration and from A. inguinalis by longer note duration. Overall, the call of A. chorus is most similar to that of A. laticeps in showing a more similar range in pulse repetition rate and note duration. However, amplitude modulation within notes differs among calls of both species ( Fig. 3 View FIGURE 3 ) and in A. laticeps pulse rate increases within notes, a character missing in calls of A. chorus . For comparative numerical call parameters see Table 2.
Natural history. At the type locality Ambodivoahangy, a large chorus consisting of many males called around midnight of 3 April 2010 from a large swampy area of a rice plantation at the forest edge. Remarkably, calls of the males were not emitted randomly but more or less synchronously, resulting in “waves” of calls interrupted by about 1.5 second intervals of silence. This behaviour made it extremely difficult to locate individual calling males, suggesting that an anti-predator strategy might be involved. Other anuran species calling at the same time were Blommersia variabilis , Boophis tephraeomystax , and Guibemantis cf. kathrinae . Single individuals were occasionally found on the forest floor during daytime, as documented for other populations of rainforest Aglyptodactylus .
Distribution. Currently reliably known only from the three mentioned lowland localities in the north-east of Madagascar ( Fig. 2 View FIGURE 2 ).
Etymology. The specific epithet is a Latin noun and refers to the calling behaviour of the species. At the type locality, males of the species called almost synchronized in a large chorus , resulting in a very conspicuous impression to the human ear.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |