
Aglyptodactylus inguinalis ( Günther, 1877 ), Gunther, 1877
|
publication ID |
https://doi.org/10.11646/zootaxa.4006.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:136F727A-1628-449F-B762-254371D27360 |
|
DOI |
https://doi.org/10.5281/zenodo.5684810 |
|
persistent identifier |
https://treatment.plazi.org/id/039D87F4-FFAF-C808-08C1-FF4CFDCDE3A6 |
|
treatment provided by |
Plazi |
|
scientific name |
Aglyptodactylus inguinalis ( Günther, 1877 ) |
| status |
|
Aglyptodactylus inguinalis ( Günther, 1877) View in CoL —bona species
Remarks. This species has been referred to as Aglyptodactylus sp. aff. madagascariensis ”Ranomafana” by Glaw & Vences (2007) and Aglyptodactylus sp. 3 by Vieites et al. (2009) and Wollenberg et al. (2011), and as Aglyptodactylus sp. Ca3 by Perl et al. (2014).
Material examined. BMNH 1947.2.26.54, holotype (probably a male, currently unavailable), from Anzahamaru, [central eastern] Madagascar; ZSM 5213/2005 ( FGZC 2802), adult female, from Marojejy, 14°26.260' S, 49°46.533' E, 481 m a.s.l., northern Madagascar; ZSM 183/2002 ( FGMV 2001.1482), adult male, from Tolagnaro, southeastern Madagascar; ZSM 2271/2007 ( ZCMV 5429), adult female, from Manombo Special Reserve (precise coordinates not taken, but roughly at 23°02' S, 47°44' E, 30‒40 m a.s.l.), southeastern Madagascar.
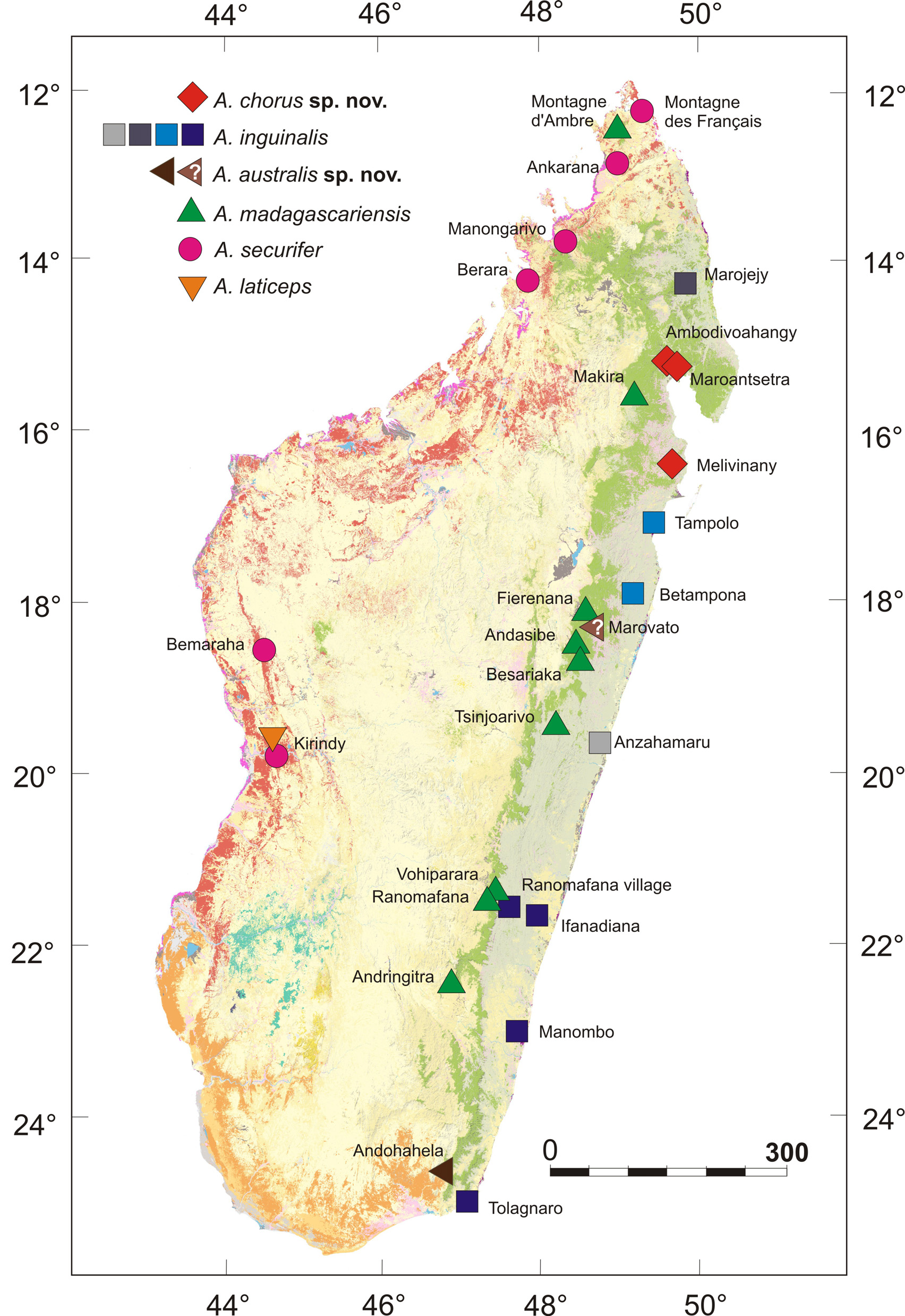
Identity. Described as Rana inguinalis from Anzahamaru by Günther (1877) and first considered to represent a junior synonym of A. madagascariensis by Boulenger (1918). According to Günther (1877), the type locality Anzahamaru most probably corresponds to a village close to Mahanoro, south of Tamatave (= Toamasina). This locality was later suggested to correspond to Anjahamaro, a locality close to Mahanoro by Brygoo (1990). Later, Blommers-Schlösser & Blanc (1991) by mistake called this locality Anzahamani in their Aglyptodactylus distribution map and placed it at the east coast, but considerably north of Toamasina. According to the information provided in the literature, we here consider Anzahamaru (= Anjahamaro) to most probably represent a locality very close to Mahanoro.
All samples from this area of the east coast corresponded to the coastal clade, with two strongly divergent sublineages, one from the Betampona/Tampolo area and one from south-eastern localities. It might be speculated that the Anzahamaru population probably belongs to the Betampona/Tampolo sub-lineage since these three sites are located north of the Mangoro river estuary which is known to act as a barrier to dispersal for some coastal frog lineages (e.g., Gehring et al. 2012). The comparatively small size of the holotype (probably a male; Table 3) agrees well with other specimens of the coastal clade (e.g., ZSM 183/2002 from Pic St. Louis near Tolagnaro), and its relative head length (0.398) fits well with the values of other coastal specimens (see section on morphological differentiation above). We therefore assign this nomen to the populations of the coastal clade. Due to the strong differentiation among the three sub-lineages here assigned to A. inguinalis (Marojejy vs. Tampolo/Betampona vs. south-east) it cannot be excluded that this is indeed a species complex warranting further subdivision, in which case it would be necessary to ascertain the identity of the Anzahamaru population by new collections in this area.
Morphology of reference specimen. ZSM 183/2002, adult male ( Fig. 11 View FIGURE 11 a, b), as indicated by brown nuptial pads on the first finger. Intercalary element between ultimate and penultimate phalanges presumably present given the typical bent-down pattern of toe tips. Body slender; head slightly broader than body; snout pointed in dorsal view, nostrils directed dorsolaterally, distinctly nearer to snout tip than to eye; canthus rostralis concave, loreal region distinctly concave, tympanum distinct, tympanum round, its horizontal diameter 63% of eye diameter; distinct supratympanic fold; tongue oval bifid posteriorly. Vomerine teeth distinct in two well separated, round patches, the diameter of the group of vomerine teeth about the same diameter as choanae. Arms slender, fingers with distinct single subarticular tubercles, outer metacarpal tubercle oval, slightly elevated; inner metacarpal tubercle prominent and elliptical; fingers without webbing; comparative finger length 2=4<1<3, slightly developed terminal finger disks, without circummarginal grooves; limbs slender, tibiotarsal articulation reaches widely beyond snout tip if limb is adpressed along body; foot with an elongated inner metatarsal tubercle, no recognizable outer metatarsal tubercle, subarticular tubercle distinct, single, rounded; terminal toe disks slightly developed, lateral metatarsalia deeply separated, comparative toe length 1<2<3<5<4. Webbing formula 1(1), 2i (1.5), 2e (0.5), 3i (2), 3e (1), 4i (2), 4e (1.5), 5 (0.5). Skin on the upper surface smooth, throat smooth, chest and anterior throat tip with keratinized spicules. Venter slightly granular and covered with similar but smaller keratinized spicules. For morphological measurements see Table 3.
Colour of reference specimen ( ZSM 183/2002) in preservative (after 13 years in alcohol): head, dorsum, and dorsal surfaces of limbs brown. Inguinal region with a small and sharply bordered black marking on each side. Flanks greyish. Loreal and temporal region greyish, tympanum brown. A fine black line below the supratympanic fold and a triangular blotch posterior to tympanum. A blackish line running from the anterior margin of the eye to tip of snout, expanding to triangular shape at mid-level between eye and nostril. Upper lip whitish. Dorsal surface of hands whitish covered with minute brown spotting, dorsal surface of feet cream; thighs and shanks with brown crossbands. Posterior surfaces of thighs without a reticulated pattern. Cloacal region dark brown. Ventral surfaces of feet dark brown. Arms greyish-brown with some irregular brown markings. Iris blackish, pupil grey. Belly and throat cream except for the brown spicules, anterior venter and ventral surface of arms cream.
In life, the general pattern was the same as in preservative, except for brown dorsal surfaces exhibiting a yellowish tint ( Fig. 11 View FIGURE 11 a) and the venter and throat being bright yellow with brown spicules ( Fig. 11 View FIGURE 11 b).
Variation. For variation in morphological measurements see Table 3. There is a considerable amount of variation in colour pattern among specimens collected throughout the huge supposed range of this species. This mainly refers to the extent and distribution of dark markings, flecks and spots on flanks and blotches and stripes in the canthal and tympanal regions of the head. These vary from being very few, faint and indistinct to numerous and/or dark and sharply outlined. However, in life, brown dorsal markings seem to be rather indistinct in most specimens. In some specimens, the iris is golden with some dark brown reticulation in both, upper and lower iris area ( Fig. 11 View FIGURE 11 a), whereas in other specimens the lower iris area can be distinctly darker ( Fig. 11 View FIGURE 11 c). Given the genetic differentiation within A. inguinalis and the small number of voucher specimens examined, more morphological variation than currently documented can be expected.
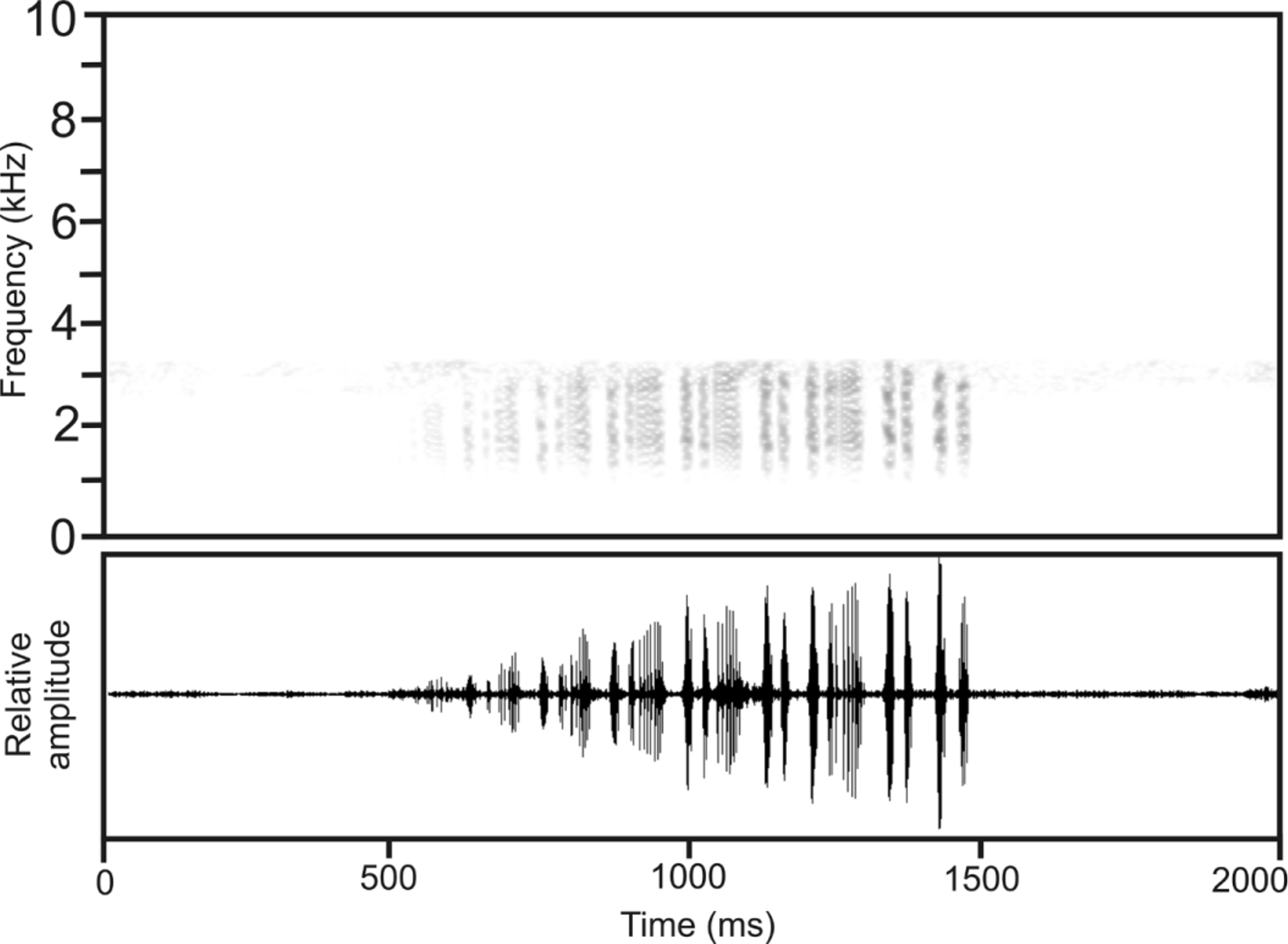
Call. The call of A. inguinalis , recorded at Tolagnaro, differs remarkably from calls of other Aglyptodactylus species in being composed of short and longer notes combined in a somewhat irregular pattern. A typical call is shown in Figure 12 View FIGURE 12 , showing the following alternation of notes: short, long, short, long, short, short, long, short, short, long, short, short, short, long, short, short, short, short. This note pattern is similar but not identical in all calls and may also vary slightly among calls of the same individual. All calls show overall amplitude modulation with almost constantly increasing call energy from the beginning to the end of the call. Both short and long notes are pulsed, with distinctly higher pulse rate in short notes. Number of pulses per note is very similar in short and long notes, and note duration seems to result from modulation of the pulse repetition rate. In some calls, pulses within short notes are indistinct or partly fused. For numerical call parameters refer to Table 2.
The release call of Aglyptodactylus madagascariensis from Betampona provided by Rosa et al. (2011) probably corresponds to A. inguinalis , given the sequence data provided by Rosa et al. (2012) and included herein.
Natural history. To our knowledge, there are no available data particularly referring to populations assigned to this species.
Distribution. As defined here based on genetics, bioacoustics and type information, we tentatively consider the following populations to represent A. inguinalis: Anzahamaru ( type locality), Betampona, Ifanadiana, Manombo, Marojejy, Ranomafana village, Tampolo, and Tolagnaro ( Fig. 2 View FIGURE 2 ). These localities are all located along the east coast of Madagascar at elevations from sea level up to 481 m a.s.l. (highest locality Marojejy at a campsite locally known as "Camp Mantella ").

| ZSM |
Bavarian State Collection of Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |