
Archaeopteryx siemensii, DAMES, 1897
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2006.00245.x |
|
persistent identifier |
https://treatment.plazi.org/id/039E8901-FFE1-9C72-FF0B-058BB07281A5 |
|
treatment provided by |
Felipe |
|
scientific name |
Archaeopteryx siemensii |
| status |
|
ARCHAEOPTERYX SIEMENSII DAMES, 1897
Referred specimen
WDC-CSG-100, deposited in the Wyoming Dinosaur Center, Thermopolis, WY, USA .
Measurements
See Table 1.
Description and comparison
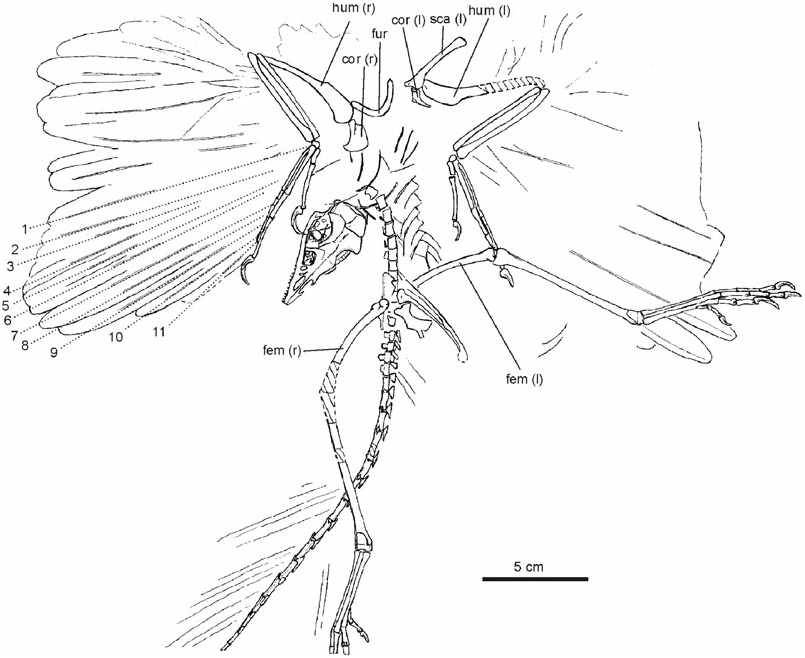
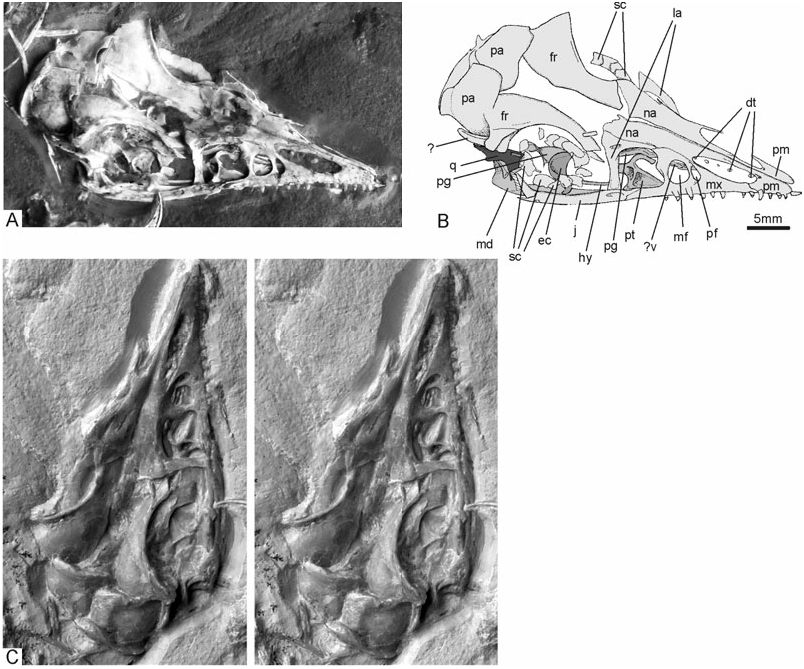
Skull: The skull ( Fig. 4 View Figure 4 ) is the only archaeopterygid skull exposed in dorsal view, allowing the recognition of some previously unknown osteological details. For example, contrary to all other avians, the premaxillary bones of Archaeopteryx are not co-ossified, not even in their most distal part, as in the enantiornithine Gobipteryx ( Chiappe, Norell & Clark, 2001) . In fact, in the new specimen, the distal ends of the premaxillae are not even in contact, so that there is a small notch at the tip of the snout. This may be an artefact of preservation, but it is notable that there is also a notch on the tip of the rostrum in the early *After Wellnhofer (1993).
†After El{anowski (2001b).
Cretaceous Confuciusornithidae ( Chiappe et al., 1999) , whose premaxillae are fused, however. The virtually uncompressed snout forms a steep roof with a subtriangular cross-section; dorsally the premaxillary bones meet at an angle of about 85°. The nasal openings are elongate and measure nearly one-sixth of the length of the skull; there is a small foramen at their dorsodistal margin. The maxillare exhibits a nearly circular maxillary fenestra and a smaller, more distal, promaxillary fenestra ( Figs 4 View Figure 4 , 5 View Figure 5 ), most likely indicating pneumatization of the antorbital sinus ( Witmer, 1990). These fenestrae occur in other theropods, but are much larger in Archaeopteryx than in, for example, dromaeosaurs ( Norell & Makovicky, 2004: fig. 10.4).
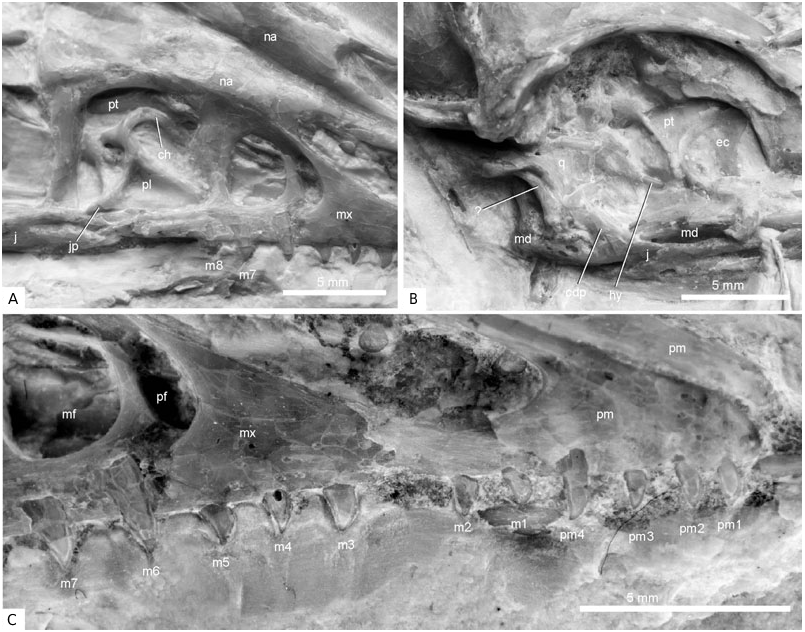
Four premaxillary and eight maxillary teeth are visible; as indicated by a gap between the second and third preserved maxillary teeth, one maxillary tooth may be missing ( Fig. 5 View Figure 5 ). An isolated tooth of uncertain provenance lies close to the tip of the right premaxilla. As in the other specimens of Archaeopteryx , the teeth show some variation in size and shape ( Wellnhofer, 1992: fig. 19). The first and second premaxillary teeth are more peg-like, whereas most others have convex rostral and concave caudal margins, and a slightly caudally projecting tip. The fourth premaxillary tooth is the longest tooth, the eighth maxillary tooth the shortest. As in other archaeopterygid specimens, none of the teeth are serrated.
The lacrimale is an ‘L’-shaped bone with a pronounced nasal process; a prefrontal appears to be absent (the bone identified as a prefrontal in the Eichstätt specimen by Wellnhofer, 1974 has been considered as part of the lacrimale by subsequent workers, for example, El{anowski & Wellnhofer, 1996; Paul, 2002). The orbital margin of the frontal forms a low rim. Owing to the fact that the brain cavity is collapsed, the frontal bones are not in medial contact. The caudal margins of the frontals are distorted against the frontal margins of the parietals, resulting in the misleading impression of a ‘temporal cavity’ on the right side of the skull, caudal to the orbits. Five scleral ossicles are preserved in articulation in the left orbit, about 13 sclerotic plates (or fragments thereof) can be counted in the right orbit.
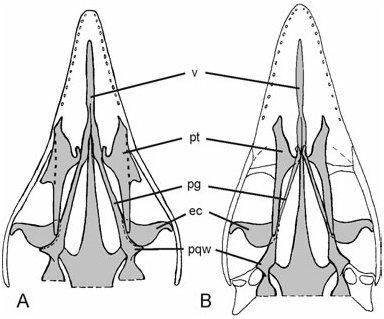
The palate of Archaeopteryx was poorly known until Wellnhofer (1993) and El{anowski & Wellnhofer (1996) described the isolated palatal elements of the Munich specimen. The new skeleton provides further anatomical details of the shape and orientation of the archaeopterygid palatine, ectopterygoid, and pterygoid ( Figs 4–6 View Figure 4 View Figure 5 View Figure 6 ). The dorsal surface of the midsection of the right palatine is visible through the antorbital fenestra, whereas most of the pterygoid wing and maxillary process are hidden by overlying sediment and bones. The bone is tetraradiate as in non-avian theropods, and the short jugal process, reported for the first time ( Mayr et al., 2005), contacts the jugal. The seemingly triradiate palatine of the Munich specimen exhibits a breakage line along its lateral margin (El{anowski & Wellnhofer, 1996: fig. 4B), and a small associated fragment (El{anowski & Wellnhofer, 1996: fig. 4E) may represent the broken jugal process. As in the Munich specimen, the dorsal surface of the palatine exhibits two marked depressions separated by a transverse crest. A small fracture in the bony wall of the deeper caudal depression indicates that it was hollow and thus probably pneumatic. As assumed by El{anowski (2001a), the hook-shaped choanal process touches the rostral portion of the pterygoid. Contrary to El{anowski’s (2001a) reconstruction ( Fig. 6B View Figure 6 ), however, the lateral margin of the maxillary process contacts the jugal and maxillare. This difference from El{anowski’s (2001a) otherwise excellent reconstruction is a result of the fact that the palatine of Archaeopteryx has a jugal process and is thus wider than assumed by El{anowski (2001a), and because the distal part of the skull appears to have been narrower than in El{anowski’s reconstruction ( Fig. 6 View Figure 6 ).
In the dorsal section of the maxillary fenestra, an elongated element can be discerned which we identify as part of the vomer (see El{anowski & Wellnhofer, 1996; El{anowski, 2001a). Also observable in the ventral portion of the maxillary fenestra is an osseous structure which may be part of the palatal shelf of the maxillare.
The ectopterygoid, which appears to be homologous to the avian uncinatum (El{anowski, 1999), is preserved in its original position, with the convex margin directed caudally. Its shape corresponds to the ectopterygoid of the Munich specimen (El{anowski & Wellnhofer, 1996: fig. 4F). Its position in the new specimen agrees with that reconstructed by El{anowski (2001a), except that the caudomedial margin tightly joins, and slightly overlaps, the rostral end of the prequadrate wing (El{anowski, 2002; triangular wing of El{anowski & Wellnhofer, 1996) of the right pterygoid, whereas it contacts the rostral blade of the pterygoid alone in El{anowski’s (2001a) reconstruction ( Fig. 6 View Figure 6 ). The hook-shaped jugal process meets the jugal.
The pterygoid also resembles the corresponding element of the Munich Archaeopteryx and confirms the hypothesized orientation of this element by El{anowski & Wellnhofer (1996) and El{anowski (2001a). In WDC-CSG-100, the caudal part and the rostral blade, which contacts the choanal process of the palatine (El{anowski, 2001a), are visible, whereas the intervening section is hidden under overlying bones. The portion caudal to the prequadrate wing is markedly twisted. The new specimen confirms the interpretation by El{anowski & Wellnhofer (1996) that the prequadrate wing is directed laterally, but its caudal margin, which was identified as the articulation facet for the quadrate by El{anowski & Wellnhofer (1996), does not contact the quadrate in the new specimen. We thus conclude that the prequadrate wing of the pterygoid of Archaeopteryx braced the ectopterygoid and did not contact the quadrate ( Fig. 6 View Figure 6 ), as in other non-avian theropods.
The temporal region of WDC-CSG-100 ( Fig. 5 View Figure 5 ) is difficult to interpret and is apparently not completely preserved in the specimen, as neither a squamosal nor a postorbital can be discerned. These bones are shown to be present in the Munich specimen, in which a squamosal with a process for articulation with the postorbital can be identified (El{anowski & Wellnhofer, 1996: fig. 6B). We assume that these bones were lost when the slab was split. A structure that, at first glance, resembles the dorsal temporal bar ( Fig. 4 View Figure 4 ), whose presence in Archaeopteryx has been controversial (El{anowski, 2001a; Paul, 2002), is either a rib, as other ribs are preserved in close proximity, or the dorsal margin of the right prootic (unfortunately, the depth of this structure cannot be discerned). There is a well-developed occipital crest caudally adjacent to the parietal bones.
The caudal end of the jugal bifurcates into a slender quadratojugal and a marked caudodorsal process, as reconstructed by El{anowski (2001a). We could not, however, identify the quadratojugal itself, which, in Archaeopteryx , is a small, ‘L’-shaped bone (El{anowski & Wellnhofer, 1996).
A substantial portion of the right quadrate seems to be preserved, although this bone appears to be damaged, making the identification of osteological details impossible. An osseous bar just caudal to the presumed caudal margin of the quadrate has so far defied identification.
Most of the mandible is hidden under the skull. Only the caudal part of the right ramus is visible caudal to the jugal and continues above the jugal. The tips of five dentary teeth, presumably from the left mandibular ramus, can be seen through the right nasal opening.
There is a thin osseous bar in the right orbit, above the right mandibular ramus, which we consider to be part of the hyoid ( Figs 4 View Figure 4 , 5 View Figure 5 ). The caudal part of this bar is covered by plates of the sclerotic ring, but apparently continues lateral of the prequadrate wing of the pterygoid and terminates on the right quadrate.
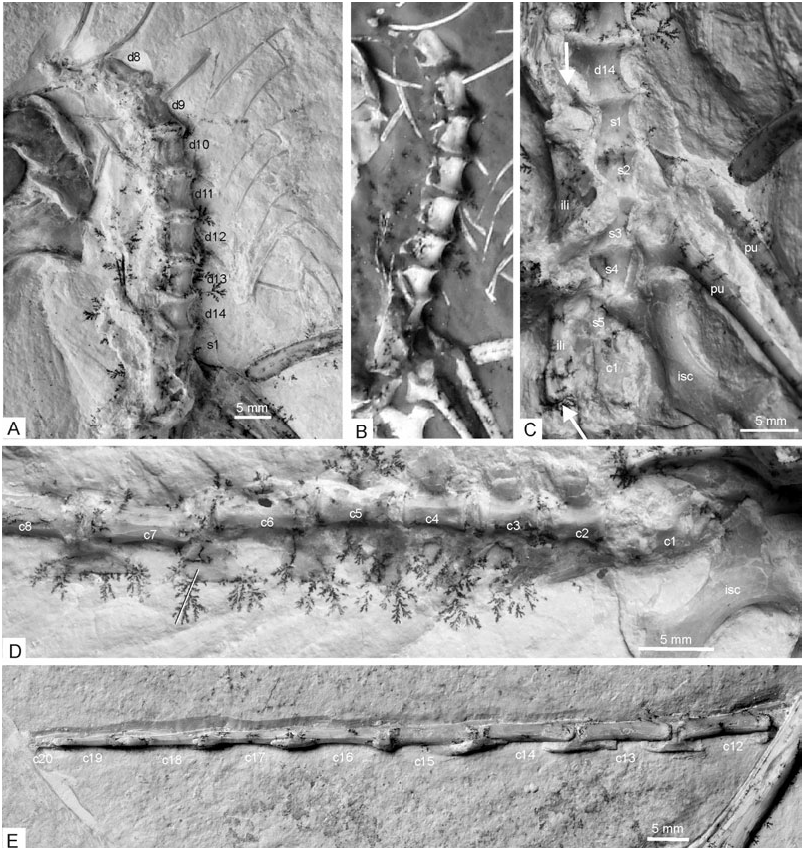
Vertebrae: The morphology of the vertebral column of Archaeopteryx is still rather poorly known. In the new specimen, three incompletely prepared cervical vertebrae in articulation are situated underneath the right manus; three caudally adjacent ones, also in articulation, are still hidden in the sediment and only visible on the X-ray photograph ( Fig. 7 View Figure 7 ). These six cervical vertebrae are not in articulation with either the thoracic vertebrae or the skull, and only few osteological details can be discerned on the X-ray photograph. They are visible in ventral view and increase in length with increasing distance from the dorsal vertebrae. The vertebra closest to the dorsal series is only slightly longer than wide, whereas the most cranial one is nearly twice as long as wide. The cranial ends bear well-developed transverse processes.
The centra of the seven most caudal dorsal vertebrae, from the thoracosacral series, are well preserved ( Fig. 8 View Figure 8 ). These are exposed in ventrolateral view and have convex lateral and ventral surfaces. Contrary to other Mesozoic non-neornithine birds, e.g. Confuciusornithidae and Ichthyornithidae ( Peters, 1996; Chiappe et al., 1999; Clarke, 2004), they do not bear marked lateral excavations (‘pleurocoels’), but only shallow, elongate depressions. As in the basal dromaeosaur Microraptor ( Hwang, Norell & Gao, 2002) , the preserved dorsal vertebrae do not exhibit pneumatic foramina, whose presence has been observed for some cervical and cervicothoracic vertebrae of Archaeopteryx ( Britt et al., 1998) , and also appear solid on the X-ray photograph. The second and third dorsal vertebrae exhibit small concave parapophyses at the cranial end of the centrum, which are slightly elevated, but not set on small stalks as appears to be the case in Microraptor ( Hwang et al., 2002) and Confuciusornis ( Chiappe et al., 1999) . Transverse processes cannot be discerned. As in the basal avian Rahonavis ( Forster et al., 1998) , there is, however, a marked laterally directing infradiapophyseal fossa at the base of the neural arches (partly filled with sediment in the specimen). Other details of the neural arches are not visible.
Given the presumed number of 23 presacral vertebrae in Archaeopteryx (El{anowski, 2002), ten presacral vertebrae are missing or at least not visible in the new specimen (there appear to be additional vertebrae beneath the cranium, but this cannot be clearly discerned on the X-ray photograph).
We assume that five vertebrae are fused into a synsacrum ( Fig. 8 View Figure 8 ) as in other specimens of the Archaeopterygidae (El{anowski, 2002), although the transition between the dorsal and sacral vertebrae is difficult to ascertain (the vertebra which is here identified as the caudalmost dorsal vertebra, ‘d14’, in Fig. 8 View Figure 8 also appears to be tightly sutured to the caudally adjacent one, here identified as the cranialmost synsacral vertebra; in this case, six vertebrae would be fused into the synsacrum, as in Rahonavis, Forster et al., 1998 ). The boundaries between the centra can still be discerned.
Twenty tail vertebrae are preserved (the distal half of the 20th is missing) and, given the number of 21–22 tail vertebrae in other Archaeopterygidae ( Wellnhofer, 1974; El{anowski, 2002), the distalmost one or two vertebrae seem to be missing ( Fig. 8 View Figure 8 ). The second to fourth tail vertebrae bear well-developed transverse processes, the fifth a small one. These vertebrae also bear large, plate-like chevrons, which become much lower and more elongated towards the distal tail vertebrae. The chevrons at the tip of the tail are very small. The tail is twisted along its longitudinal axis, i.e. the proximal third is seen in ventrolateral view, the midsection in lateral view, whereas the distal section, beginning with the 16th caudal vertebra, exposes its ventral surface. Thus, it can be discerned that the 17th−19th tail vertebrae are strongly mediolaterally compressed, with very narrow centra (the width of the 17th centrum is only 0.7 mm). The lengths of the centra of the tail vertebrae are (in mm): 3.8 (second), 4.2 (third), 4.2 (fourth), 5.4 (fifth), 6.5 (sixth), 7.9 (seventh), 9.5 (eighth), ∼10.0 (?) (tenth), 11.1 (11th), 10.9 (12th), 10.9 (13th), 10.6 (14th), 10.6 (15th), 10.1 (16th), 9.1 (17th), 9.1 (18th).
Ribs and gastralia: Only a few ribs are preserved in the specimen, owing to the fact that the thoracic vertebrae proper are lacking. Some ribs are damaged, so that it can be discerned that they were hollow. For the first time, the gastralia are seen in ventral view ( Fig. 8 View Figure 8 ). Their arrangement is not symmetrical, because the elements form a zigzag pattern as in the Confuciusornithidae ( Chiappe et al., 1999: fig. 33). Of the seven caudal rows, elements from both sides are preserved and each row apparently consists of four elements; on each side, two of these overlap for nearly half of their length; of the four cranialmost rows, only the right elements are visible.
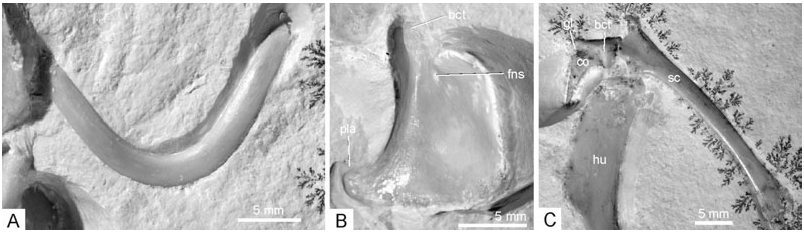
Coracoid: The morphology of the coracoid of Archaeopteryx has been controversial, owing to the fact that none of the hitherto known specimens shows a complete view of this bone (compare the reconstructions in, for example, de Beer, 1954; Ostrom, 1976; Martin, 1991; Wellnhofer & Tischlinger, 2004). In the new specimen, nearly the entire right coracoid is exposed in cranial view and, in addition, the lateral surface of the left one can be seen ( Fig. 9 View Figure 9 ). The body of the bone is of subrectangular shape, with a concave lateral margin and a well-developed lateral process. Except for the more pronounced lateral process, it agrees well in its shape with the coracoid of the basal avian Sapeornis as reconstructed by Zhou & Zhang (2003a: fig. 6f), whereas the coracoid of other basal avians, such as Jeholornis and Confuciusornis ( Chiappe et al., 1999; Zhou & Zhang, 2003b), is narrower and more elongated. The distal (sternal) margin is slightly convex. As already noted by de Beer (1954), the bone is bent craniocaudally along a line running obliquely from the biceps tubercle to the edge corresponding to the angulus medialis ( Baumel & Witmer, 1993) of the coracoid of extant birds. The biceps tubercle (preglenoid process of El{anowski, 2002) is elongated. In lateral view, the bone resembles the coracoid of the basal dromaeosaur Sinornithosaurus ( Xu, Wang & Wu, 1999) . It widens towards the glenoid extremity, the sternal extremity is narrow; the area between the biceps tubercle and the glenoid is concave. There appears to be a small foramen for the supracoracoideus nerve ( Fig. 9 View Figure 9 ). However, although this foramen is preserved in the expected position and appears to be real, there are similar foramina near the medial margin of the bone which seem to be preparation artefacts. The medial margin does not exhibit any incisions, which were assumed by de Beer (1954: fig. 4) for the London specimen.
Scapula: The left scapula is exposed in lateral aspect ( Fig. 9 View Figure 9 ); the right scapula is still hidden in the matrix, but visible on the X-ray photograph ( Fig. 10 View Figure 10 ). The bone is much shorter than the humerus, the left scapula measures 35.0 mm. The acromion is narrow, the surface between it and the glenoid fossa concave. The corpus of the scapula widens slightly toward its truncate caudal end (contra Martin, 1985: fig. 3). Apart from the narrower acromion, the scapula of Archaeopteryx is similar in morphology to that of Sinornithosaurus ( Xu et al., 1999) . The left coracoid and scapula join at an angle of about 90°, but, as in all other Archaeopterygidae with the possible exception of the London specimen ( de Beer, 1954), are not fused.
Furcula: The boomerang-shaped furcula ( Fig. 9 View Figure 9 ) has its cranial surface exposed, and is similar in shape to that of the London specimen. The bone is curved craniocaudally, with the midsection of the scapi clavicularum protruding most strongly cranially. As in the Confuciusornithidae ( Peters, 1996; Chiappe et al., 1999), the extremitas omalis is simple and rounded. An acromial process is not developed. The furcula appears to have been only loosely connected to the other pectoral bones, which may explain its absence in the Eichstätt and Berlin specimens.
Sternum: In concordance with other specimens of the Archaeopterygidae , there are no ossified sternal plates. The alleged presence of an ossified sternum in the Munich specimen has recently been disproved ( Wellnhofer & Tischlinger, 2004).
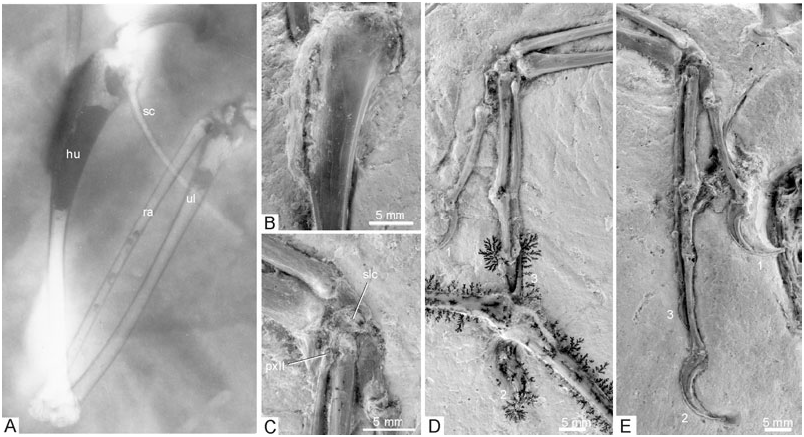
Humerus: The cranial surface of the right humerus is exposed ( Fig. 10 View Figure 10 ), whereas the left one is seen from its caudodorsal side (the distal half of this bone has been restored by the preparator). In contrast with modern birds but as in the ninth specimen and non-avian theropods ( Wellnhofer & Röper, 2005), the main plane of the distal end is twisted at an angle of about 45° against that of the proximal end. The proximal end of the humerus lacks any of the fossae and sulci characteristic of extant birds, and the caput humeri is directed more ventrally than in extant birds. The crista deltopectoralis measures slightly more than one-third of the entire length of the bone. In concordance with other Mesozoic avians (e.g. Confuciusornithidae , Jeholornis ), it projects laterally without any cranial deflection known in extant birds. The caudal surface of the humerus is flat, the cranial surface medial of the crista deltopectoralis slightly convex. There is a small crista bicipitalis. The distal end of the right humerus remains embedded in the sediment, but is discernible on the X-ray photograph ( Fig. 10 View Figure 10 ). Its shape appears similar to the distal humerus of the basal avian Sapeornis ( Zhou & Zhang, 2003a: fig. 7), with the condylus ventralis protruding furthest distally and the condylus dorsalis being orientated more obliquely to the longitudinal axis of the humerus than in neornithine birds.
Ulna and radius: Both ulnae have their dorsal surfaces exposed and are preserved complete and uncrushed. The shaft is very slender in its midsection, but widens towards the distal end; it is only slightly wider than the shaft of the radius, whereas the ulna is much wider than the radius in Microraptor ( Hwang et al., 2002; Xu et al., 2003). An olecranon is virtually absent, as is a well-developed processus cotylaris dorsalis. In contrast with the basal avian Rahonavis ( Forster et al., 1998) , even faint papillae remigales cannot be discerned.
The distal end of the radius exhibits a pointed tuberculum aponeurosis ventralis ( Baumel & Witmer, 1993). Details of the proximal ends of both radii cannot be observed.
Carpalia and metacarpals: Only the semilunate carpal bone is visible in the specimen ( Fig. 10 View Figure 10 ; three other carpal bones may be visible on the ultraviolet-induced fluorescence photograph of the left wing, but their identification is uncertain). It mainly caps the second metacarpal and only the cranial edge contacts the first metacarpal, whereas ‘it caps all of metacarpal I and part of metacarpal II’ in Microraptor ( Hwang et al., 2002: 19) . The metacarpals are seen from their dorsal side. As in the Eichstätt specimen, the proximal end of the third metacarpal lies above (dorsal to) the proximal end of the second metacarpal; it is situated further distally than the proximal end of the second metacarpal (pro El{anowski, 2002; contra Gishlick, 2001). The second and third fingers are tightly joined, but in contrast with the Berlin, Eichstätt, and Solnhofen specimens, as well as the ninth one, they do not cross ( Fig. 10 View Figure 10 ). Also in contrast with these specimens, the second phalanx of the third finger is only slightly shorter than the first ( Table 1). The third phalanx of the third finger is very thin, much thinner than the second phalanx, indicating a considerable degree of reduction of this finger.
The keratinous sheaths of the ungual phalanges are well preserved on the first and second right manual digits. As in the other specimens, they show no sign of wear ( Peters & Görgner, 1992). Also as in most other specimens, their concave surface is directed cranially.
Pelvis: The elements of the pelvis are disarticulated and have moved from their original position ( Fig. 11 View Figure 11 ). Few details of the poorly preserved right ilium can be discerned; the preacetabular part has a concave lateral surface, the tapering postacetabular part is seen in ventral view and is mediolaterally wide. The right ischium is visible in lateral view and exhibits the characteristic derived archaeopterygid shape. The proximal end bifurcates into two processes for articulation with the ilium and pubis, respectively. The ventral margin is concave, the dorsal margin bears a process in its midsection (intermediate process of El{anowski, 2002; posterior process of Hwang et al., 2002). The distal end bifurcates into two processes that were termed dorsodistal and ventrodistal processes by El{anowski (2002) (the latter corresponds to the obturator process of non-avian theropods). The narrow dorsodistal process is separated from the wider ventrodistal one by a deep incision. In its shape, the ischium corresponds to that of the Eichstätt and Munich specimens as reconstructed by Wellnhofer (1974, 1993). It is dorsoventrally wider than the ischium of A. lithographica as reconstructed by El{anowski (2002: fig. 6.4), and there are no proximodorsal process and foramen obturatum. The ischium of Archaeopteryx resembles the corresponding bone of Microraptor ( Hwang et al., 2002) and, to a lesser degree, the basal troodontid Sinovenator ( Xu et al., 2002) , in which the distal end is not bifurcated by a notch between the two processes.
There is a marked pit on the lateral surface of the expanded proximal end of the right pubis. As in the London and Eichstätt specimens (El{anowski, 2002), the pubic boot is partially replaced by a calcite mass, possibly indicating the former presence of cartilage. Proximal to the pubic boot, the pubic apron extends over almost half of the length of the pubis.
Femur: The right femur is seen in medial view (its distal portion has been restored by the preparator), the left one from its craniomedial side. The shaft of the bone is craniocaudally curved. The femoral head is orientated nearly perpendicular to the longitudinal axis of the bone. On the cranial surface of the proximal end, there is a depression between the lesser trochanter and the femoral head. As in other basal avians (e.g. Confuciusornithidae, Chiappe et al., 1999 ) and in Microraptor ( Hwang et al., 2002) , but contrary to modern birds, the cranial surface of the distal end lacks a patellar sulcus ( Fig. 11 View Figure 11 ).
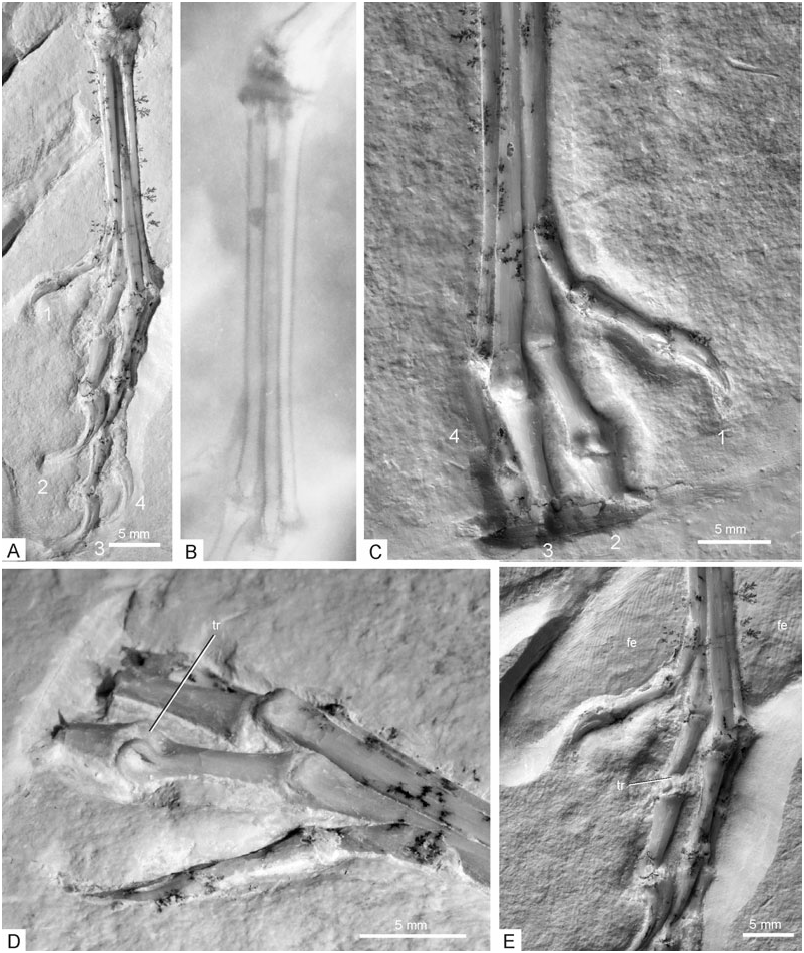
Tibia and tarsal bones: The right tibia is seen in cranial view, the left in craniomedial view ( Fig. 12 View Figure 12 ); the proximal part of the right tibia has been restored by the preparator. The proximal end of the tibia is similar to that of Microraptor ( Hwang et al., 2002) , with a well-developed cnemial crest that slants laterally. The cranial surface of the tibia is slightly convex over most of its length, whereas it is essentially flat in its distalmost section. For the first time, the proximal tarsals are clearly visible in the new specimen ( Mayr et al., 2005). The condylar portion of the astragalus and the calcaneus are proximodistally low, only about half the size of the proximal tarsals of the dromaeosaurs Microraptor ( Hwang et al., 2002: fig. 28) and Sinornithosaurus ( Xu et al., 1999: fig. 4). The calcaneus is very narrow, reaching only the width of the distal end of the fibula; the astragalus and calcaneus are separated by a narrow furrow, but may have been at least partly fused. As in other theropods, the astragalus forms a broad, 5.6-mm-long ascending process, which covers most of the cranial surface of the distal tibia. The ascending process is separated from the condylar portion of the astragalus by a transverse groove. The condyles of the astragalus are separated by a broad and shallow concavity, the medial condyle is more pronounced than the lateral one. On the medial side, there is a marked furrow between the distal part of the medial margin of the ascending process of the astragalus and the tibia. The distal end of the tibia further protrudes medially beyond the astragalus as in ornithomimosaurs ( Barsbold & Osmólska, 1990) and the basal avian Jeholornis ( Zhou & Zhang, 2002: fig. 3). Except for the latter feature, which appears to have not been recognized by previous authors, the tarsus of Archaeopteryx agrees well with the restoration of Paul (2002: fig. 10.14), but is very different from that of, for example, Martin (1991: fig. 43). The thin fibula widens distally just before it contacts the calcaneus. Distal tarsals cannot be discerned ( Fig. 12 View Figure 12 ).
Metatarsals: The three-dimensionally preserved metatarsals are seen in dorsal view and appear to be unfused over their entire lengths ( Fig. 13 View Figure 13 ). In their morphology and arrangement, the metatarsals compare well with those of Microraptor ( Hwang et al., 2002) . The third metatarsal is the longest and lies slightly dorsal to the second and fourth in the distal half of the metatarsus, but slightly ventral to them in the proximal half. The proximal ends of the second and fourth metatarsals are mediolaterally and dorsally expanded, whereas the third metatarsal becomes narrower in its proximal half and is proximally pinched by the second and fourth metatarsals. The foot of Archaeopteryx thus approaches the arctometatarsalian condition ( Holtz, 1995), although, in the typical arctometatarsalian foot, the proximal end of the third metatarsal is much more compressed. The distal section of the third metatarsal is slightly widened, but subapically exhibits a shallow concavity on its medial side, on the level of the distal end of the second metatarsal. The distal end of the second metatarsal is displaced further plantar than that of the third and fourth metatarsals. Mediolaterally, the second metatarsal is approximately twice as wide as the fourth and extends slightly less far distally; the proximal section of the shaft has a subrectangular cross-section. The fourth metatarsal is very narrow mediolaterally, but its distal end may have been expanded dorsoventrally. The distal articular surfaces of the metatarsals are smooth and lack the ginglymoid condition (according to Hwang et al., 2002; the articular surface of the second metatarsal is ginglymoid in Microraptor ). The distal ends of the second and third metatarsals have an essentially flat dorsal surface, whereas that of the fourth metatarsal is raised to a small point. A fifth metatarsal is not visible, but is expected on the plantar surface of the metatarsals ( Wellnhofer, 1992). In its proportions, the metatarsus of WDC-CSG-100 corresponds to that of the Berlin and Munich specimens. It is more slender than that of A. lithographica as exemplified by the London, Solnhofen, and Maxberg specimens. The ratio of maximum length to minimum width at midsection is 9.1 for the metatarsus of the new specimen and 6.2 for that of the Solnhofen specimen (after Wellnhofer, 1992: fig. 16).
Toes: As detailed by Mayr et al. (2005), and also assumed by Middleton (2002), the first toe of Archaeopteryx was not fully reversed as in modern birds (contra, for example, Wellnhofer, 1993). In the new specimen, it is spread medially on both feet ( Fig. 13 View Figure 13 ). Metatarsal I attaches to the medial (not medioplantar, contra El{anowski, 2002) side of the second metatarsal, in approximately its distal quarter, whereas it attaches to the plantar surface of the tarsometatarsus in modern birds with a fully reversed hallux ( Middleton, 2001); its proximal section even protrudes slightly further dorsad than the second metatarsal ( Fig. 13 View Figure 13 ). Moreover, the shaft does not exhibit the torsion characteristic for birds with a fully reversed hallux ( Middleton, 2001). As in Confuciusornis ( Peters, 1996; Chiappe et al., 1999), the articulation of the first toe is ball-like and medially protruding. Mayr et al. (2005) detailed that the hallux is also medially directed in the Solnhofen and Berlin specimens, in which the first toe is preserved/visible on one foot only. In the Solnhofen specimen, the metatarsals are seen from their plantar side and the proximal phalanx of the first toe from its medioplantar side; in the Berlin specimen, much of the dorsal aspect of the first toe is visible, whereas the metatarsus is seen from its medial side. In both cases, the respective aspect of the first toe would not be visible if it was fully reversed. The feet of the London and Eichstätt specimens are preserved in lateral or medial view, and the impression of a reversed first toe in these specimens may thus be an artefact of preservation, with the medially spread toe being brought level with the sedimentation layer via compaction ( Mayr et al., 2005).
The trochlea of the proximal phalanx of the second toe is proximodorsally expanded ( Fig. 13 View Figure 13 ), indicating that the toe was hyperextendible, as in dromaeosaurs, troodontids, and the basal avian Rahonavis ( Mayr et al., 2005) . According to current phylogenies (e.g. Gauthier, 1986; Sereno, 1999; Hwang et al., 2002; Xu et al., 2002; Benton, 2004), this feature must be regarded as a synapomorphy of Paraves, i.e. a clade ( Troodontidae + Dromaeosauridae + Aves) that is lost in birds more derived than Archaeopteryx and Rahonavis . Just proximal to the expanded trochlea there is a marked depression on the dorsal surface of the proximal phalanx of the second toe. Such a depression is also present on the distal end of the proximal phalanx of the third toe which lacks, however, a dorsally protruding articulation. The depressions of the insertion area of the collateral ligaments are marked. The flexor tubercles of the ungual pedal phalanges are weak. The fourth toe consists of five phalanges, but there are only four in the Solnhofen specimen. The keratinous sheaths of the ungual phalanges of the second and fourth digits of the left foot are well preserved.
Feathers: The specimen exhibits well-preserved wing and tail feather impressions. Impressions of the barbs are best visible in the distal portion of the primaries; barbules cannot be discerned. On the right wing, the rachises of 11 primaries can be counted ( Fig. 3 View Figure 3 ). The most distal (11th) primary is the shortest, the fourth the longest. The approximate lengths of the primaries are as follows (in mm; the minimum lengths of some primaries of the Berlin specimen, from Rietschel, 1985, are given in parentheses): 118 (first; 130), 123 (second), 125 (third; 135), 133 (fourth), 129 (fifth; 145), 125 (sixth), 120 (seventh; 140), 117 (eighth), 106 (ninth; 125), and 87 (tenth; 95). The length of the 11th primary cannot be measured, but appears to have been at least 47 mm (some barbs are preserved in that area). Three primaries insert on the second phalanx of the second digit, two on the first, and the remaining six on the metacarpal ( Fig. 3 View Figure 3 ). The same insertion pattern was inferred by Rietschel (1985) for the Berlin specimen. The distalmost primary apparently inserts in the midsection of the second phalanx, which indicates that the ungual phalanx of the second digit was not covered by feathers and was therefore functional. At least the eighth to tenth primaries have an asymmetric vane.
In the Berlin and London specimens, the odd-numbered rachises are only preserved as ‘shaft shadows’ without impressions of the vanes ( Rietschel, 1985; El{anowski, 2002). This led some previous workers to assume that the primaries of Archaeopteryx were arranged in two different levels (El{anowski, 2002). However, in the new specimen, a distinction between shafts and shadow shafts cannot be made, and at least the four most distal primaries (eighth to 11th) exhibit true shaft impressions.
The exact number of secondaries cannot be counted, but may have been about 12–15 as assumed for the Berlin specimen ( Stephan, 1987).
Many rachises exhibit a furrow along their midline which, by comparison with modern birds, indicates that, as in the Berlin specimen ( Heinroth, 1923), their ventral surfaces are exposed, whereas, as noted above, the forearm skeleton is seen from its dorsal side (see Helms, 1982 for an explanation of this kind of preservation). Curiously, however, in the distal part of the right wing, the outer vanes overlap the inner ones of the distally adjacent feathers, as is characteristic of the dorsal aspect of a modern birds’ wing. Especially in the proximal area of the secondaries of the left wing, marked furrows can be discerned, which measure up to 50 mm and run obliquely to the longitudinal axis of the secondaries, at an angle of about 30°. We assume that these are impressions of the coverts, by comparison with modern birds probably the lower ones which run more obliquely than the dorsal coverts. It thus appears likely that impressions of the ventral surfaces of the wings are preserved.
Although the impressions of the primaries are rather faint, there are marked, fuzzy furrows at the ‘elbow joint’ which may stem from the tertiaries. Similar furrows of uncertain identity can also be found next to the proximal end of the right humerus and below the left tibia.
The number of preserved tail feathers cannot be counted, as both the rachises and the feather margins left an impression in the sediment. Again, impressions of barbs are visible, although they are fainter than those in the wings. All tail feathers meet the vertebrae at the same angle of about 30°. The length of those inserting on the 11th and 12th caudal vertebrae is about 65 mm, whereas the feather attaching to the 14th vertebra measures about 75 mm. The impressions of the feathers attached to the proximal seven tail vertebrae are more irregular than the feathers attached to more distal vertebrae.
Impressions of hindlimb feathers cannot be discerned (see Christiansen & Bonde, 2004 for their presumed presence in the Berlin specimen).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Archaeopteryx siemensii
| Mayr, Gerald, Pohl, Burkhard, Hartman, Scott & Peters, D. Stefan 2007 |
Archaeopteryx
| VON MEYER 1861 |
Archaeopteryx
| VON MEYER 1861 |