
Nematocarcinus burukovskyi, Komai & Segonzac, 2005
|
publication ID |
https://doi.org/ 10.5281/zenodo.5401000 |
|
persistent identifier |
https://treatment.plazi.org/id/039F210B-6A60-BC3A-FF7E-4A63EBD0FA9B |
|
treatment provided by |
Marcus |
|
scientific name |
Nematocarcinus burukovskyi |
| status |
sp. nov. |
Nematocarcinus burukovskyi View in CoL n. sp. ( Figs 1-7 View FIG View FIG View FIG View FIG View FIG View FIG View FIG )
HOLOTYPE. — BIOSPEEDO, DS Nautile dive PL1587 , basket 1, SEPR, Garrett Segment, Wormwood site, marker BS 15, 17°34.91’S, 113°14.68’W, 2595 m, 28.IV. 2004, subadult cl 18.6 mm, tl c. 63 mm (MNHN-Na 15768). GoogleMaps
PARATYPES. — PAR5, DS Alvin, dive 4094, slurp gun, SEPR, Fred’s Fortress site, 31°09.06’S, 111°55.88’W, 2330 m, 29.III.2005, 1 young cl 13.8 mm ( CBM-ZC 8360 ) GoogleMaps ; dive 4096, slurp gun, SEPR, Serpulid Mound site, 23°32.07’S, 115°34.17’W, 2612 m, 01.IV.2005, 1 cl 25.2 mm ( USNM) GoogleMaps .
, Wormwood vent site, SEPR, cl 18.6 mm (MNHN-Na 15768),
ETYMOLOGY. — Named in honor of Rudolf N. Burukovsky, who published an essential revision of the Nematocarcinidae .
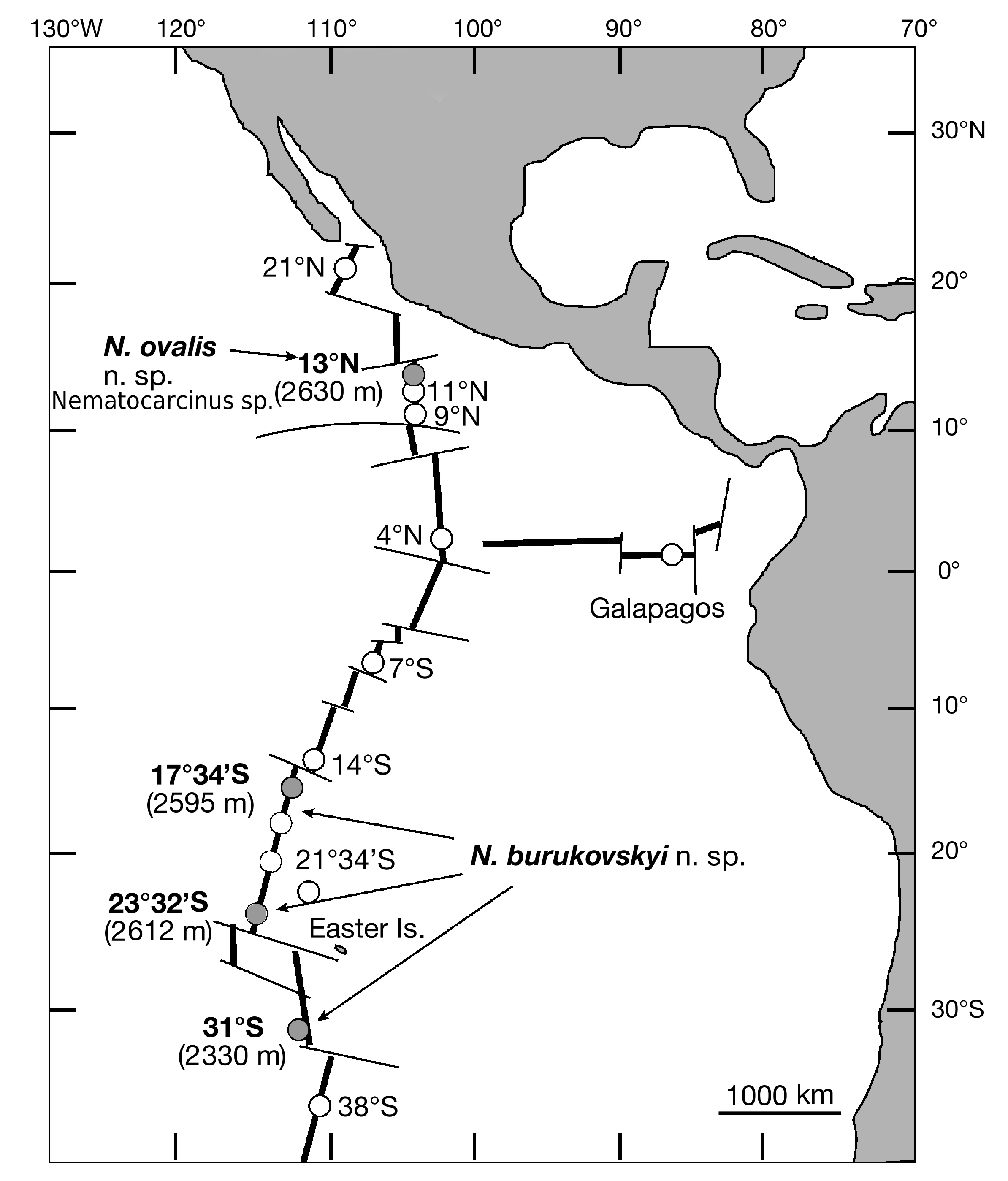
DISTRIBUTION. — Known from the South East Pacific Rise 17°S, Garrett Segment, 2595 m (type locality) and south of Easter Island at 23°S, 2612 m, and 31°S, 2330 m ( Fig. 1 View FIG ).
DESCRIPTION
Holotype
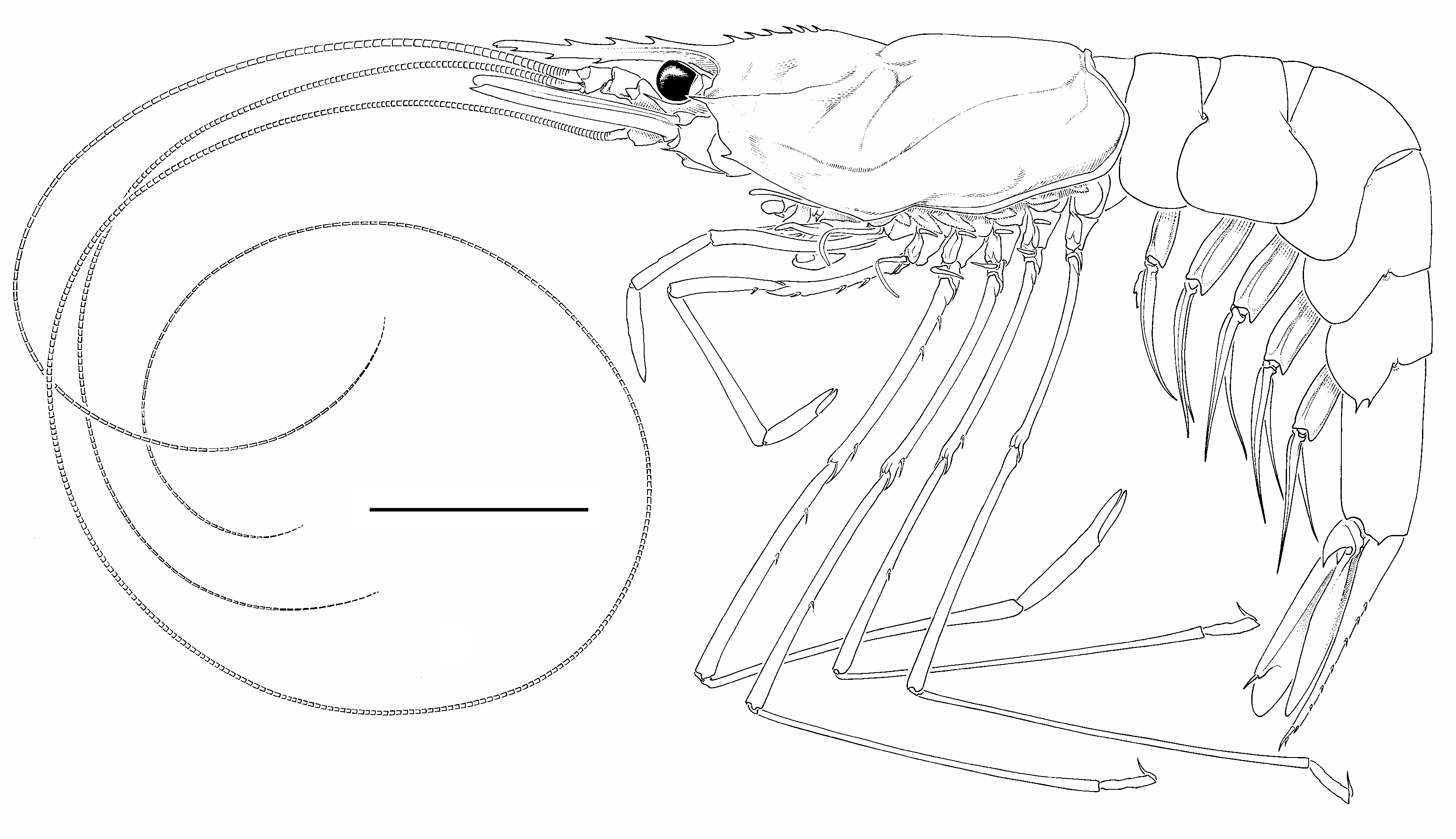
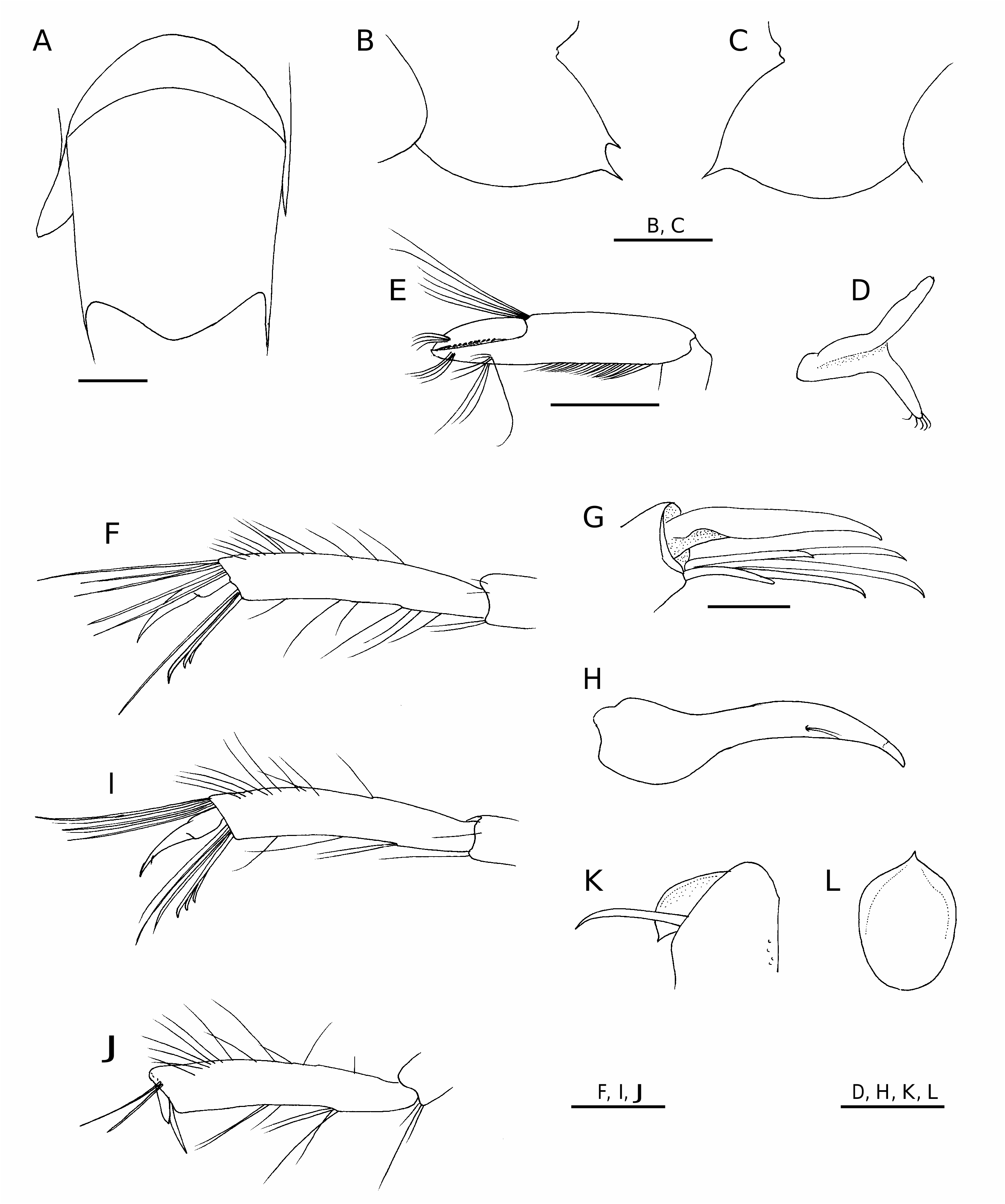
Body ( Fig. 2 View FIG ) moderately slender; integument soft, not membranous, surface smooth, shiny.
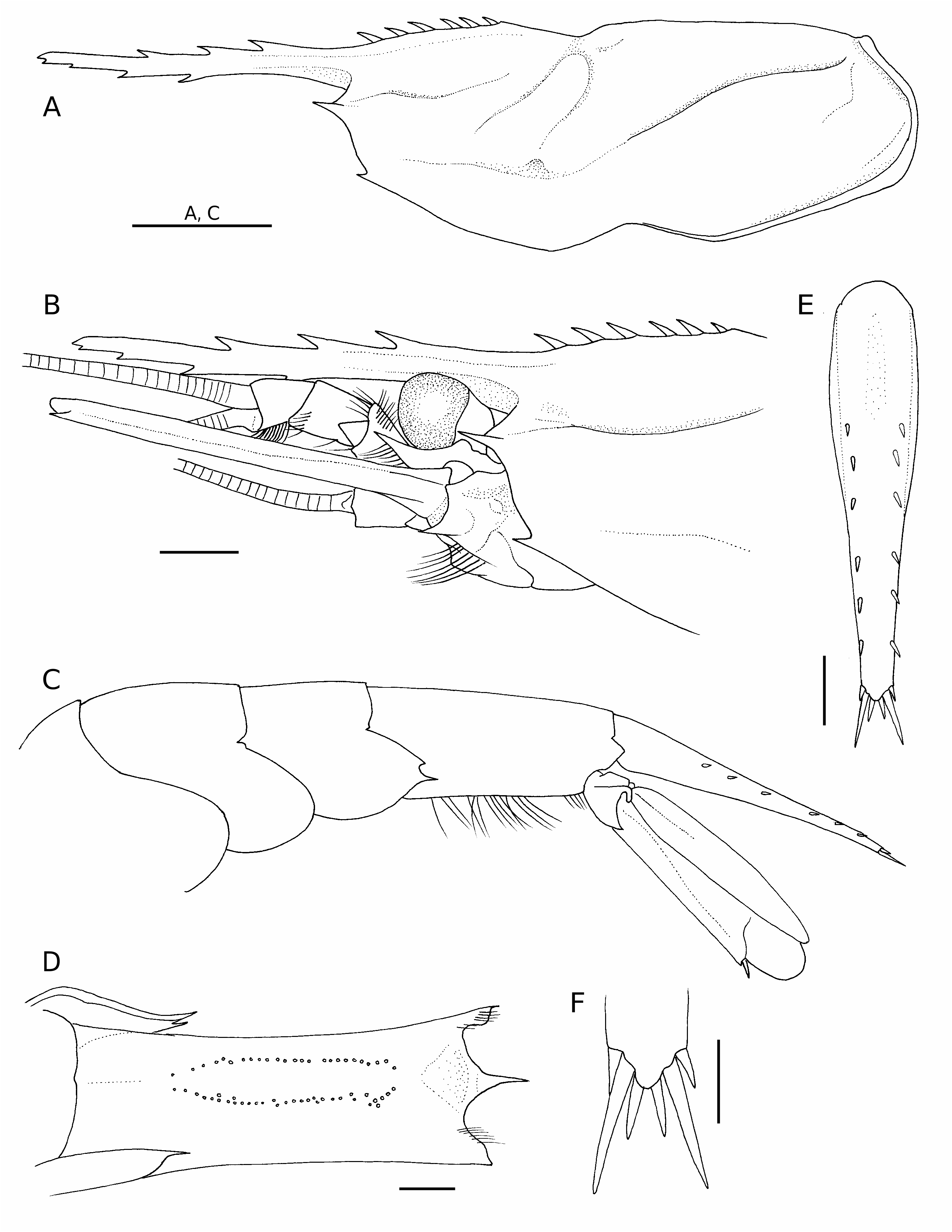
Rostrum ( Fig. 3A, B View FIG ) slender, slightly falling short of anterior margin of antennal scale, 0.62 of carapace length; dorsal margin armed with three fixed teeth, distal 0.35 unarmed; ventral margin with four fixed teeth, subterminal small; tip weakly deflexed; lateral surface with blunt lateral ridge confluent with orbital margin. Carapace ( Fig. 3A, B View FIG ) with six post-rostral, basally articulat- ed spines, most anterior spine widely separated from most posterior tooth on rostrum proper, most posterior spine arising at 0.28 of carapace length; post-rostral ridge low, sharp, extending to about 0.40 of carapace length; lateral surface well sculptured with distinct postorbital, cervical, post-cervical, hepatic, branchiocardiac grooves; cervical groove across midline of carapace, shallow but conspicuous notch on dorsal surface; branchiocardiac groove without sharply delineat- ed ridge; orbital margin evenly rounded, without suborbital lobe; antennal spine well developed, reaching level of anterior margin of basicerite, acuminate; pterygostomial spine smaller than antennal spine; anterolateral margin between antennal, pterygostomial spines oblique, slightly sinuous.
Thoracic sternum ( Fig. 4I View FIG ) with three bilobed prominences on sixth to eighth somites; lobes of prominence on sixth somite each terminating in acute spine; lobes of posterior two prominences rounded.
Abdomen ( Fig. 2 View FIG ) dorsally smooth. Pleura of anterior four somites broadly rounded. Posterodorsal margin of third somite only weakly produced, rounded posteriorly ( Fig. 6A View FIG ). Fifth somite ( Fig. 3C View FIG ) with two acute posteroventral teeth on left (dorsal tooth distinctly smaller than ventral tooth) ( Fig. 6B View FIG ), with slender posteroventral tooth on right ( Fig. 6C View FIG ); inner surface of posterior part of pleuron smooth, without tubercle or ridge-like structure ( Fig. 3D View FIG ). Sixth somite ( Fig. 3C, D View FIG ) moderately elongated, 1.94 times longer than fifth somite, 2.26 times longer than anterior depth; posterolateral process subtriangular, subacutely pointed; posteroventral angle blunt; ventral surface with single row of long setae on either side of midline, almost parallel except for anterior, posterior parts curving toward midline; paired posteroventral spots absent ( Fig. 3D View FIG ); pre-anal tooth ( Fig. 3D View FIG ) direct- ed posteriorly. Telson ( Fig. 3C, E View FIG ) moderately long, 4.70 times longer than greatest width, reaching posterior margin of exopod of uropods (except for terminal spines), gradually tapering posteriorly to blunt terminal lobe; dorsal surface with shallow median sulcus in anterior one-third, with six pairs of small dorsolateral spines; all dorsolateral spines aligned; terminal margin with three pairs of spines, second pair longest (about 3.0 of length of lateralmost pair).
Eye ( Fig. 3B View FIG ) subpyriform with well developed, faceted cornea; cornea moderately large for nematocarcinid, its width about 0.10 of carapace length; eye-stalk distinctly shorter than corneal width.
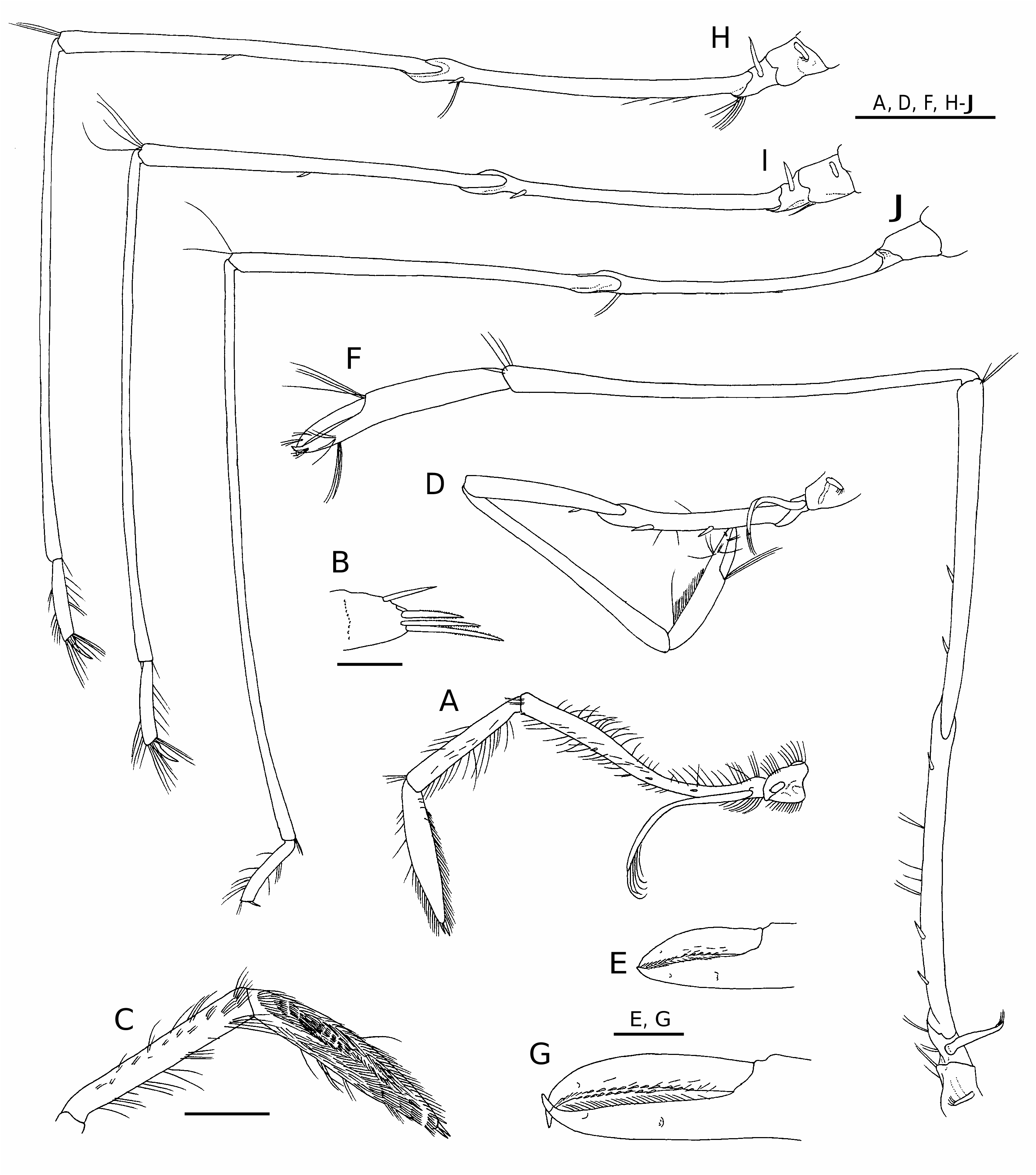
Antennular peduncle ( Fig. 3B View FIG ) stout, reaching midlength of antennal scale. First segment subequal in length to distal two segments combined; stylocerite slightly falling short of distal margin of first segment, acuminate, weakly upturned in lateral view. Flagellar length exceeding twice that of carapace length ( Fig. 1 View FIG ).
Antennal peduncle ( Figs 3B View FIG , 4H View FIG ) with basicerite stout, armed with acute ventrolateral distal tooth; ventral surface of basicerite provided with low, rounded tubercle. Fifth segment (carpocerite) short, stout, reaching distal margin of first segment of antennular peduncle. Antennal scale 0.56 of carapace length, 4.00 times longer than wide; lateral margin very slightly sinuous; distolateral tooth reaching broadly rounded lamella. Flagellum longer than body ( Fig. 1 View FIG ).
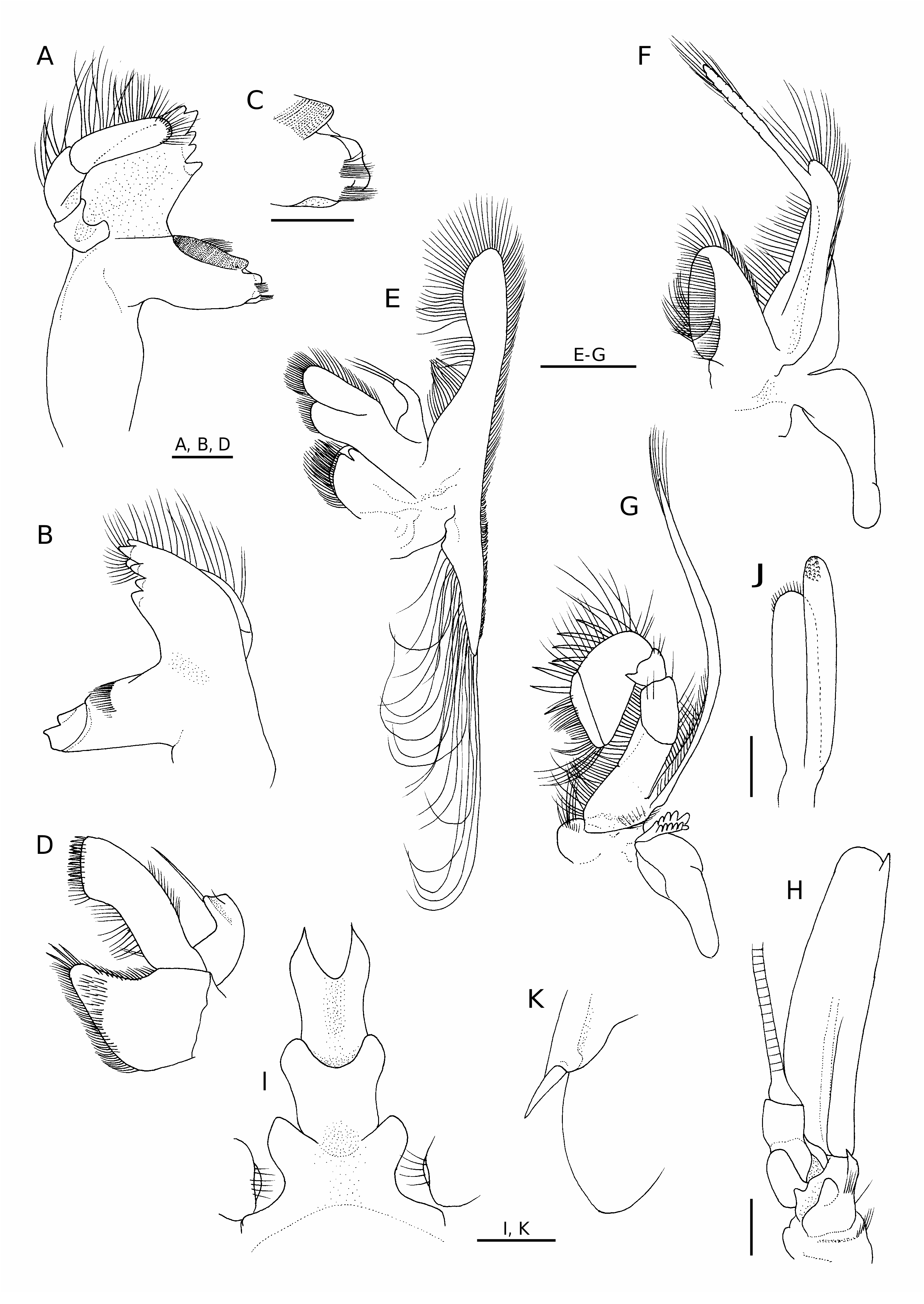
Mouthparts typical of genus. Mandible (left) ( Fig. 4 View FIG A-C) with broad incisor process terminating in five strong, corneous teeth; molar process with finely ridged, dentate distal margin; anterior surface of molar process with transverse rows of stiff setae, narrow space present between setal row, surface of molar process; mesial face of molar process obliquely truncate, ornamented with numerous rows of minute setules; inner distal angle with two tufts of short setae; palm curved mesially, consisting of three articles, distal article longer than proximal two articles combined, with numerous stiff setae on margins. Maxillule ( Fig. 4D View FIG ) with coxal endite tapering distally, with some slender spines distally, stiff short setae marginally, outer surface also with short setae in distal part; basial endite somewhat curved mesially, distal margin truncate, armed with two rows of slender spines, stiff setae; endopod strongly curved, bilobed distally, inner lobe with long apical bristle, outer lobe with short subapical seta. Maxilla ( Fig. 4E View FIG ) with coxal endite rounded, very unequally bilobed (distal lobe very small, rudimentary); mesial margin of coxal endite double edged; basial endite well exceeding coxal endite, deeply bilobed (distal lobe slightly longer than proximal lobe); endopod basally stout, strongly curved mesially, with three long apical setae; scaphognathite narrow, posterior lobe elongated triangular, with row of very long setae on mesial margin extending to tip. First maxilliped ( Fig. 4F View FIG ) with basial endite rounded subtriangular in outline; outer surface of basial endite convex, with two separate longitudinal rows of setae, each row extending proximally toward mesial margin; coxal endite broken off; endopod simple, distinctly overreaching distal margin of basial endite, with row of numerous setae on mesial margin; exopod with narrow caridean lobe, long flagellum; epipod large, deeply bilobed, posterior lobe elongated. Second maxilliped ( Fig. 4G View FIG ) with endopod strongly flexed at articulation between propodus, carpus, composed of six segments, but vestigial suture between ischium, basis discernible; dactylus narrow, with four long spines distomesially; propodus with row of four long spines on mesial margin; carpus with sharp tooth at distomesial angle; coxa somewhat produced mesially; exopodal flagellum long, slender; epipod large, with small, but distinctly lamellate podobranch. Third maxilliped ( Fig. 5A View FIG ) with endopod not reaching distal margin of antennal scale, consisting of four segments; ultimate segment somewhat compressed laterally, lance shaped, with terminal cluster of small, setulose spines ( Fig. 5B View FIG ); mesial face of ultimate segment ( Fig. 5C View FIG ) with numerous transverse or obliquely transverse tracts of short to stiff setae of various length, forming grooming apparatus; carpus (penultimate segment) subequal in length to ultimate segment, with scattered tufts of short to moderately long setae on surfaces; antepenultimate segment (merus-ischiumbasis fused segment) flattened dorsoventrally, sinuous in dorsal view, with row of four spinules on lateral surface (including spinule at ventrolateral distal angle), with prominent tufts of long setae on dorsal surface proximally; epipod consisting of rounded dorsal process situated on lateral face of coax, strap-like ventral projection, latter without terminal hook ( Fig. 6D View FIG ); exopod about 0.60 length of antepenultimate segment.
First to fifth pereopods ( Figs 1 View FIG ; 5D, F View FIG , H-J) slen- der, posterior four pereopods elongated, but degree of slenderness and elongation lesser compared with other congenerics; articulations between merus, ischium strongly oblique, typical for nematocarcinids.
First pereopod ( Figs 5D, E View FIG ; 6E View FIG ) overreaching antennal scale by length of chela; dactylus 0.70 of palm length, terminating in acute, curved claw, cutting edge with two rows of spinules, short setae; fixed finger also terminating in curved claw, crossing dactylus, with tuft of setae at midlength of lateral surface, cutting edge with two rows of short stiff setae. Palm with tuft of long setae at articulation to dactylus, patch of grooming setae on ventromesial surface. Carpus widened distally, 1.88 of chela length. Merusischium combined 1.25 of carpus length, 0.65 of carapace length; merus slightly widened distally, with one small ventrolateral spine subp r o x i m a l l y; i s c h i u m s u b e q u a l i n l e n g t h t o merus, armed with two widely spaced spines ventrolaterally. Exopod short, reaching midlength of ischium, with several short marginal setae.
Second pereopod ( Fig. 5F, G View FIG ) longest among five pairs of pereopods, overreaching antennal scale by length of chela and carpus. Chela large for genus, 0.42 of carapace length. Dactylus 0.62 of palm length, terminating in acute, curved claw, cutting edge with two rows of spinules, short setae; fixed finger also terminating in curved claw, crossing dactylus, with two tufts of setae at midlength of lateral, ventral surfaces, cutting edge with two rows of short stiff setae. Palm with tuft of long setae at articulation to dactylus; carpus elongated, widened distally, 2.15 of chela length. Merus-ischium combined 1.37 of carpus length, 1.33 of carapace length; merus slightly widened distally, with two small, widely spaced ventral spines in proximal half; ischium subequal in length to merus, armed with three widely spaced spines (one subdistal, two subproximal) ventrolaterally. Exopod very short, reaching proximal 0.20 of ischium, with few very short terminal setae.
Third to fifth pereopods similar in length and structure. Third pereopod ( Fig. 5H View FIG ) overreaching antennal scale by full length of distal three segments. Dactylus ( Fig. 6 View FIG F-H) 0.50 of propodus length, peculiar in shape, flattened dorsoventrally, abruptly narrowed at proximal one-third; proximal part with convex lateral, mesial margins, lateral margin forming thick keel; ventral surface of proximal part excavate; distal part notably curved mesially, terminating in slender, acuminate unguis, with tuft of setae on dorsal surface. Propodus ( Fig. 6F, G View FIG ) very short, subcylindrical, slightly widened distally, with row of individual or tufts of setae on dorsal, ventral surfaces; distal margin obliquely truncate, with row of long stiff setae dorsally and row of five or six slightly curved spines, some of them longer than dactylus. Carpus 6.24 of propodus length. Merusischium combined 1.23 of carapace length; articulation between merus and ischium notably inflated; merus with one ventral spine at midlength, ischium with one ventrolateral spine subdistally. Exopod short. Fourth pereopod ( Figs 5I View FIG ; 6I View FIG ) overreaching antennal scale by 0.85 of carpus length; dactylus 0.40 of propodus length; merus-ischium combined 1.22 of carapace length; exopod rudimentary, shorter than that of third pereopod. Fifth pereopod ( Fig. 5J View FIG ) overreaching antennal scale by 0.30 of carpus length; dactylus ( Fig. 6 View FIG J-L) very short, flattened, strongly appressed to obliquely truncate distal margin of propodus, rounded in dorsal (extensor) view, terminating in small acute point; margins of dactylus sharply edged; propodus ( Fig. 6J, K View FIG ) generally similar to those of third, fourth pereopods (most spines on ventrodistal margin missing); carpus 7.8 of propodus length; merus-ischium combined 1.24 of carapace length, both segments unarmed; exopod absent.
Gill formula typical of genus, summarized in Table 1. Epipods on first to fourth pereopods all strap-like, but non-hooked ( Fig 5A, D, F, H, I View FIG ).
First pleopod with endopod elongated oval, with small appendix interna arising at distal 0.25. Second to fifth pleopods ( Fig. 1 View FIG ) each with well developed appendix interna. Appendix masculina of second pleopod ( Fig. 4J View FIG ) shorter than appendix interna, narrow, with row of very short setae on rounded terminal margin. Exopods moderately long, that of third pleopod longest, 0.42 of carapace length. Uropod ( Fig. 3C View FIG ) with stout protopod terminating posterolaterally in acute tooth; exopod slightly longer than endopod, with distinct suture; lateral margin of exopod straight, terminating in very small, acute tooth, long movable spine arising just mesial to posterolateral tooth ( Fig. 4K View FIG ).
Notes on paratypes
T h e t w o p a r a t y p e s, a f e m a l e (c l 2 5.2 m m; USNM) and a juvenile of undetermined sex (cl 13.8 mm; CBM-ZC 8360), agree well with the holotype. Rostrum (partly broken in female paratype) armed with three dorsal and four ventral teeth; distal 0.40 of dorsal margin unarmed; tip weakly deflexed as in holotype. Carapace with six (young specimen) or seven (female) postrostral teeth, only posterior four with basal suture in female, all with basal suture in young specimen. In both specimens, left fifth abdominal pleuron with one posteroventral tooth, right with two posteroventral teeth. Sixth abdominal somite with single (young) or double (female) row of setae on ventral surface either side of midline. First pereopod with three ventrolateral spines on ischium. Second pereopod with one or two lateral spines on merus. Propodus of fifth pereopod with several long spines on ventrodistal margin.
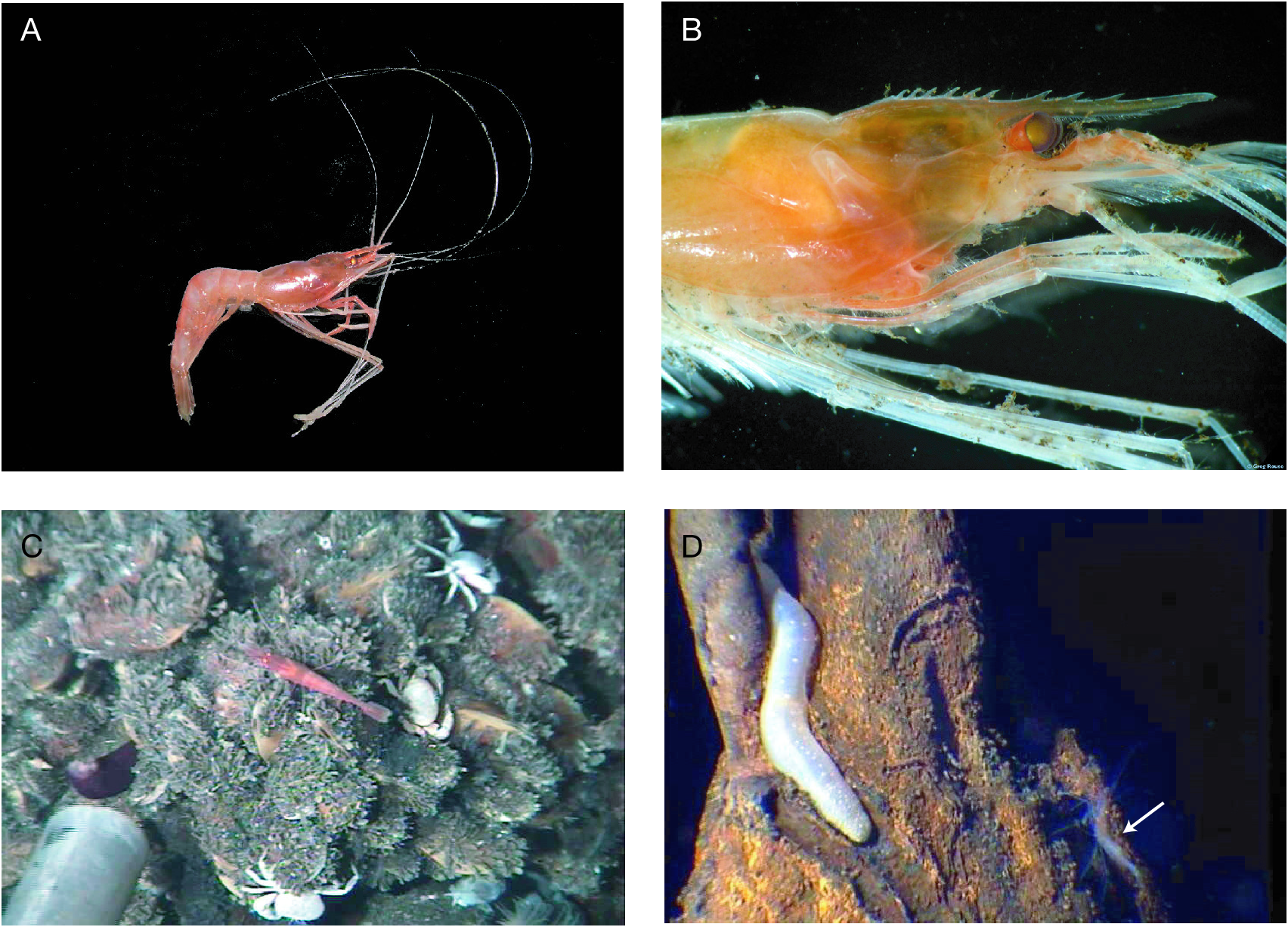
Coloration in life ( Fig. 7A, B View FIG )
Body light orange, yellowish hepatopancreas visible through integument. Cornea of eye brown, but reflective; eye-stalk dark orange. Pereopods nearly colorless.
Size
The largest specimen is a female cl 25.2 mm, tl c. 97 mm.
REMARKS
The holotype is a male, having a fairly well differentiated appendix masculina ( Fig. 4J View FIG ). Nevertheless, the appendix masculina is rather narrow and shorter than the appendix interna; the endopod of the first pleopod is devoid of grooves or ridges on the surfaces. Generally in Nematocarcinus , the appendix masculina is broad and longer than the appendix interna, and the endopod of the first pleopod has sculptured surfaces. It is thus thought that the holotype is a subadult. It was not possible to determine the sex of the smallest paratype (cl 13.8 mm), because differen- t i a t i o n o f t h e g o n o p o r e s w a s n o t e v i d e n t. Nevertheless, the three specimens are quite well consistent in every diagnostic aspects discussed below.
The new species is assigned to Nematocarcinus with some hesitation because of the existence of some particular characters. It is characterized by: 1) conspicuous cervical and branchiocardiac grooves on the carapace; 2) absence of paired posteroventral spots on the sixth abdominal somite; 3) small podobranch on the second maxilliped; 4) the dorsal teeth on the rostrum clearly separat- ed in two groups; 5) long posteroventral tooth of the fifth abdominal pleuron; 6) second pereopod is the longest among the five pereopods and has a relatively large chela; and 7) short exopods on the pereopods, in particular that on the fourth pereopod being rudimentary. The first three characters initially led us to suspect that the new species may be closely related to Nigmatullinus acanthitelsonis , because both species share these characters. Distinct cervical and branchiocardiac grooves are also seen in Nematocarcinus ovalis n. sp., N. faxoni , and N. longirostris , and therefore the development of the carapace grooves is not significant at the generic level. The presence of the paired posteroventral spots on the sixth abdominal somite needs to be verified in Nematocarcinus altus Bate, 1888 (cf. Burukovsky 2003). The shape and structure of the podobranch on the second maxilliped remains to be described for many Nematocarcinus species , although the podobranch of the species examined is larger than that of the new species. The other characters appear to be unique for the new species. In Nigmatullinus acanthitelsonis and other Nematocarcinus species , the dorsal rostral series is continuous, although the distance between the teeth may become greater distally ( Burukovsky 2003). The posteroventral tooth on the fifth abdominal somite is variable in shape, but not elongate as in N. b u r u k o v s k y i n. s p.; t h e f i f t h p l e u r o n i s unarmed in N. acanthitelsonis . The second pereopod is shorter than the posterior three pereopods, as far as known. The exopods on the pereopods are better developed, all flagellum-like. Future study may eventually reveal that N. burukovskyi n. sp. is generically distinct.
ECOLOGY
The holotype of N. burukovskyi n. sp. was collect- ed at the Wormwood vent site, Garrett Segment, SEPR (Jollivet et al. 2004), at the base of a 10 m high chimney, diffusing milky fluid, above beds of the mytilid Bathymodiolus thermophilus Kenk & Wilson, 1985 (of which shells were sometimes covered with white filamentous bacteria), close to small clumps of the giant siboglinid worm Riftia pachyptila Jones, 1981 , and among the clumps of tube worm many individuals of bythograeid crabs ( Bythograea thermydron Williams, 1980 , and B. microps de Saint Laurent, 1984 ), and galatheid squat lobsters ( Munidopsis subsquamosa Henderson, 1885 ). Ophidiid fishes, two or three octopus Vulcanoctopus cf. hydrothermalis Gonzales, Guerra, Pascual & Briand, 1998 , and clouds of amphipods also occurred at this site during the observation. At the Hobbs site (17°35.19’S, 113°14.68’W, 2595 m), about 500 m south from the Wormwood site, several individuals of nematocarcinid shrimp, probably representing N. burukovskyi n. sp., were also observed. Shrimps were swimming around the vent community of mytilid bivalves, siboglinid worms, bythograeid crabs and fish. This new species was common at the these two sites, although only the specimen designated as the holotype could be collected.
The paratype specimen collected during the PAR5 cruise (Alvin dive #4094, 31°S) was observed on mytilid mussel beds ( Bathymodiolus sp. , identified by R. von Cosel), among bythograeid crabs Bythograea spp. , some rare alvinocaridid shrimps resembling species of Chorocaris , gastropods Eosipho auzendei Warén & Bouchet, 2001 , and white sea anemones ( Fig. 7C View FIG ). The other paratype was collected during the same cruise (Alvin dive #4096, 23°S). This site, named Serpulid Mound, is characterized by an abundance of serpulid polychaete Laminatubus hydrothermica Ten Hove & Zibrowius, 1986 , otherwise shelters a depauperate vent fauna.
The photograph taken at the Krasnov vent site (21°33.59’S, 114°17.88’W, 2839 m) ( Fig. 7D View FIG ), located 7 km south of the Wormwood site (SEPR, 21°33’S, 114°17’W, 2839 m), shows a specimen presumably belonging to this new species standing close to a holothurian Chiridota hydrothermica Smirnov, Gebruk, Galkin & Shank, 2000 lying on an extinct chimney of oxidized sulphide.
The occurrence of N. burukovskyi n. sp. at the hydrothermally influenced areas indicates that it tolerates hydrothermal discharge at low temperature between 2 and 7 °C above the mussel beds (data taken during the BIOSPEEDO cruise).
| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |