
Dactylosoma kermiti, Netherlands & Cook & Du Preez & Vanhove & Brendonck & Smit, 2020
|
publication ID |
https://doi.org/ 10.1016/j.ijppaw.2019.12.006 |
|
publication LSID |
lsid:zoobank.org:pub:F5CBE7E7-D57F-4854-A98B-A6D6ED6FD342 |
|
DOI |
https://doi.org/10.5281/zenodo.11085330 |
|
persistent identifier |
https://treatment.plazi.org/id/039F87C1-FF9C-FF9B-954E-F9392761FCDA |
|
treatment provided by |
Felipe |
|
scientific name |
Dactylosoma kermiti |
| status |
n. sp. |
3.1.1. Dactylosoma kermiti Netherlands, Cook and Smit n. sp.
Type-host: Ptychadena anchietae (Bocage, 1868) ( Anura : Ptychadenidae ).
Other host: Sclerophrys gutturalis (Power, 1927) ( Anura : Bufonidae ).
Site in host: Peripheral blood.
Definitive host and vector: Unconfirmed. Observed possible vectors (see below).
Type-locality ( P. anchietae ): Ndumo Game Reserve , KZN, South Africa (S26.926179 ̊, E32.332416 ̊). GoogleMaps
Other localities ( Scl. gutturalis ): Sodwana , KZN, South Africa (S27.488591 ̊, E32.664259 ̊) GoogleMaps .
Type-material: Hapantotype, 1 × blood smear with a parasitaemia of 5.7% from Pty. anchietae deposited in the protozoan collection of the National Museum, Bloemfontein, South Africa, under accession number [ NMB P 534]. Parahapantotype, 1 × blood smear: Pty. anchietae , parasitaemia of 2.0%, deposited in the protozoan collection of the National Museum, Bloemfontein, South Africa, under accession number [ NMB P 535].
Additional material: 2 × blood smears: Scl. gutturalis with a parasitaemia of 0.2%, respectivly, deposited in the Protozoan Collection of the National Museum, Bloemfontein, South Africa, under accession number [ NMB P 536 - 537].
Representative DNA sequences: The sequence data specifically associated with D. kermiti (upon which the present biological description is based) have been submitted to GenBank and are as follows: Nuclear 18S rDNA (nu 18S) partial sequence: MN 879389 – MN 879398.
ZooBank registration: The Life Science Identifier ( LSID) of the article is urn:lsid:zoobank.org:pub:F5CBE7E7-D57F-4854-A98B-A6D6ED6FD342 .
The LSID for Dactylosoma kermiti n. sp. is urn:lsid:zoobank.org:act:0D6D6A74-4136-4A9D-AE15-73FDFE12EBD3 .
Etymology: The epithet is constructed in a positive form of a personal name. As the type host for this species is a frog, the species is named after the Muppet character, Kermit the Frog. Gender is male.
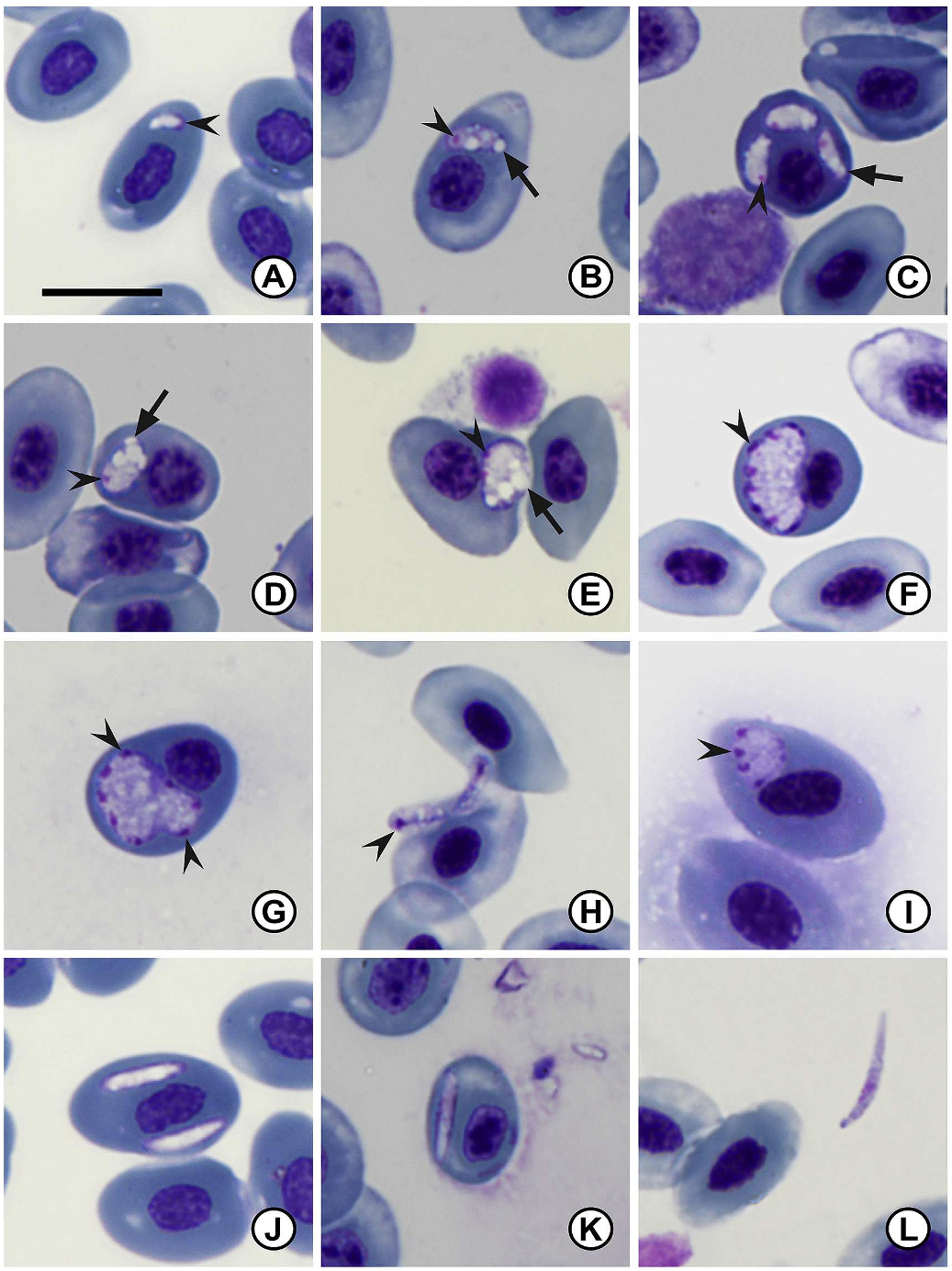
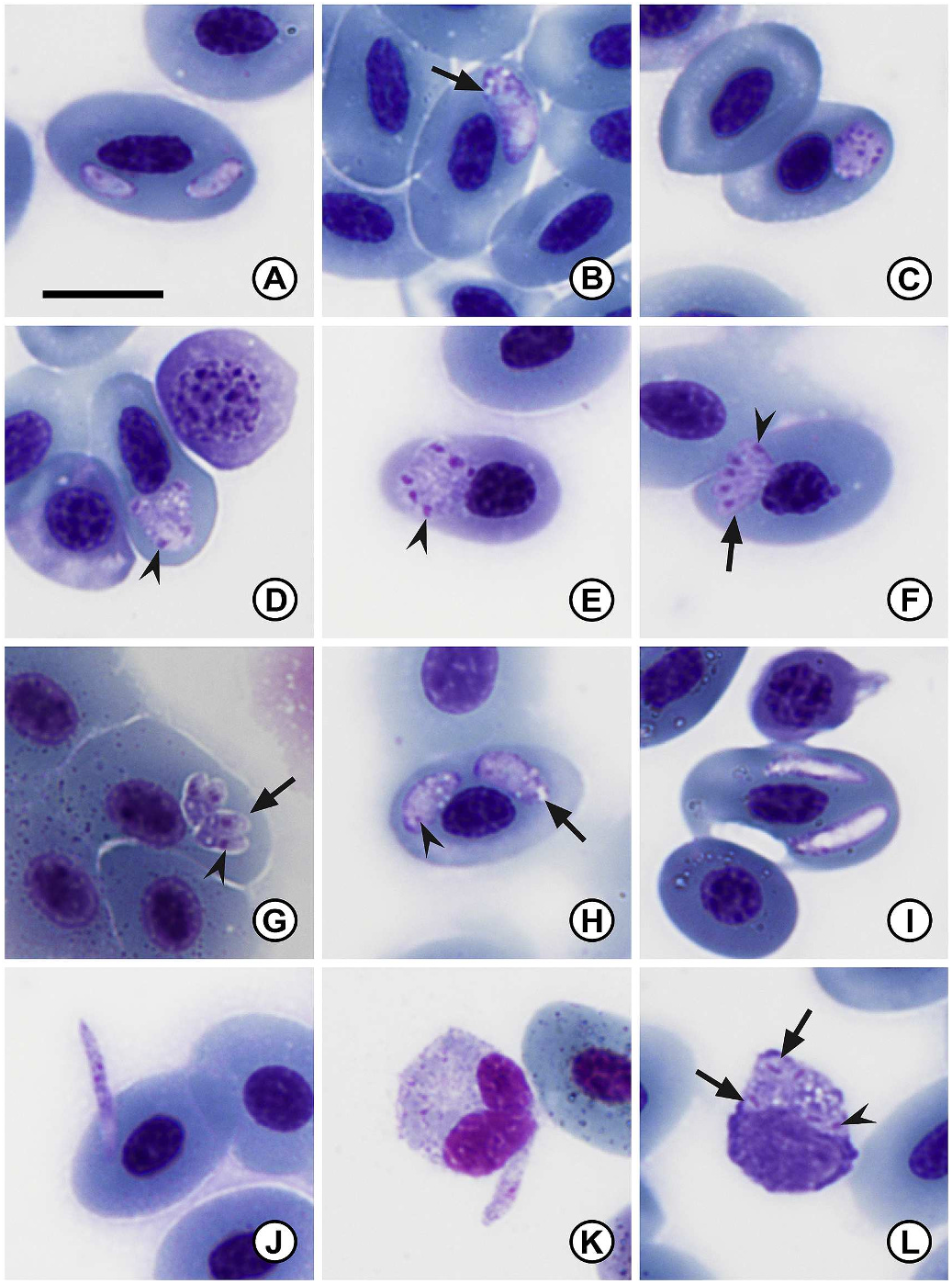
3.1.1.1. Description. Two distinct stages of merogony present in erythrocytes and occasionally in thrombocytes and monocytes of both infected type host Pty. anchietae ( Fig. 1 A–L View Fig ) and other host Scl. gutturalis ( Fig. 2 A–L View Fig ). The first stage characterised by large meronts producing up to approximately 12 merozoites. The second stage meront, seemingly smaller, produces up to six merozoites. The general appearance of the cytoplasm is hyaline, yet coarse. Parasitaemia of infected individuals (n = 70) given in percentage (%) was 0.4 ± 1.2 (0.1–5.7). The distribution range in the type host Pty. anchietae was larger with infected individuals collected from various localities and habitats, as compared to Scl. gutturalis that were only found infected at Sodwana Bay (Supplementary data S1).
Primary merogony: young trophozoites ( Figs. 1 A View Fig , 2 A View Fig ), rare, observed within erythrocytes, ovoid crescent shape, largely non-stained except peripherally. Nuclei not clearly defined, located at blunt end (broader end of the parasite). Condensed chromatin staining deep magenta ( Fig. 1 A View Fig arrowhead). Measuring 4.2 ± 0.6 (3.6–4.9) long × 2.9 ± 0.8 (1.8–3.7) wide (n = 4).
Trophozoites ( Fig. 1 B–D View Fig , 2 B View Fig ) elongated to oval, usually tapering towards one end, distinct vacuoles frequently present ( Fig. 1 B–D View Fig , 2 B View Fig arrow). Small round dense nuclei located at blunt end, chromatin staining deep magenta ( Fig. 1 B–D View Fig ). Trophozoites measure 6.7 ± 2.2 (5.3–7.7) long × 3.5 ± 1.2 (2.6–4.4) wide (n = 20).
Young primary meronts ( Figs. 1E View Fig and 2C View Fig ) large ovoid to round shape, causing slight displacement of the host cell nucleus, distinct vacuoles frequently present ( Fig. 1 E View Fig arrow). Multinucleate, nuclei division located peripherally, chromatin staining bright deep magenta ( Fig. 1 E View Fig arrowhead). Young meronts measure 7.8 ± 0.9 (6.3–9.5) long × 5.7 ± 1.3 (4.0–8.3) wide (n = 20).
Primary meronts ( Fig. 1 F, G View Fig ) varying in form, round to crescent shape, causing slight distortion or displacement of the host cell or nucleus, cytoplasm staining whitish-purple. Multinucleate, normally showing more than 10 nuclei located peripherally ( Fig. 1 F, G View Fig arrowhead), chromatin staining deep magenta. Meronts measure 9.9 ± 1.2 (8.3–12.2) long × 6.9 ± 0.8 (5.1–8.0) wide (n = 10).
Primary merozoites ( Fig. 1 H View Fig ) elongate to ovoid, hyaline cytoplasm staining pinkish-purple. Small round dense nuclei located at blunt end, chromatin staining deep magenta ( Fig. 1 H View Fig , arrowhead). Merozoites measure 5.6 ± 0.5 (5.0–6.6) long × 2.7 ± 0.4 (1.8–3.2) wide (n = 16).
Secondary merogony: young secondary meront ( Fig. 2 D View Fig ) irregular ovoid to round shape, cytoplasm staining whitish-purple, distorting the host cell and slight displacement of the host cell nucleus. Multinucleate secondary meront, nuclei located peripherally, chromatin staining bright pink to dark purple ( Fig. 2 D View Fig , arrowhead). Young meronts measure 5.4 ± 0.5 (4.3–6.3) long × 4.6 ± 0.6 (3.4–5.8) wide (n = 19).
Secondary meronts ( Figs. 1 I View Fig , 2 E–G View Fig ) differ in form, ovoid to quadrilateral shape, mature forms have dactylate (hand-like) appearance ( Fig. 2 F, G View Fig ), cytoplasm staining dark purple. Multinucleate, normally between four and six nuclei located peripherally ( Fig. 1 I View Fig , 2E–G View Fig arrowhead), chromatin staining deep magenta. Secondary meronts measure 7.2 ± 0.8 (5.6–8.6) long × 5.7 ± 0.9 (4.4–6.9) wide (n = 16).
Secondary merozoites ( Fig. 2 F–G, L View Fig arrow) elongate to ovoid, hyaline cytoplasm staining purple. Small round dense nuclei located closer to the centre, chromatin staining deep magenta ( Fig. 2 F–G, L View Fig arrowhead). Secondary merozoites measure 4.8 ± 0.5 (4.2–5.5) long × 2.3 ± 0.5 (1.8–3.5) wide (n = 11).
Gamonts ( Fig. 1 J–L View Fig , 2 H–K View Fig ) elongate and slender, often tapering to one end. In some cases slight displacement of the host cell nucleus is visible. Immature forms largely non-staining, except peripherally - staining dark purple ( Fig. 1 J View Fig ). Intracellular mature gamont forms have a hyaline appearance, although nuclei not clearly defined, nucleoplasm conglomeration visible off centre closer to blunt side ( Fig. 1 K View Fig ). Extracellular forms often elongate with a slight curvature, nuclei visible slightly off centre, staining dark purplish-pink. Gamonts measure 10.6 ± 2.5 (7.8–15.0) long × 2.2 ± 0.3 (1.5–3.0) wide (n = 40).
3.1.1.2. Remarks. Dactylosoma kermiti n. sp. can be characterised by its slender trophozoite and small merozoite stages; the number of primary (up to 14) and secondary (up to six) merozoites produced by meronts; and the vacuolated trophozoite stages. This species can be distinguished from all currently recognised species of Dactylosoma from anuran hosts, namely D. ranarum , D. sylvatica , and D. taiwanensis , based on several morphometric and developmental characteristics. Morphologically this parasite conforms closest to D. sylvatica with measurements overlapping across trophozoite, meront, merozoite and gamont stages. However, only a single, most probably the secondary merogonic cycle is reported, with meronts only producing up to eight merozoites. In comparison the number of merozoites produced by D. kermiti n. sp. differs in primary and secondary merogony with up to 14 or six merozoites observed in the primary and secondary meronts, respectively. Although measurements do overlap, D. kermiti n. sp. seems to possess generally more slender trophozoite stages and smaller merozoite stages. Also, D. sylvatica is reported from the wood frog Lithobates sylvatica from Canada. Dactylosoma ranarum is apparently a cosmopolitan species recorded from several hosts across Europe, Central and South America, Africa, and Asia. Based on morphometric data from the original description of D. ranarum , as well as from a more recent ultrastructural study of this species’ morphology, several stages differ in size when compared to D. kermiti n. sp. (see Table 1 View Table 1 ). The most notable differences as compared to the relatively smooth to finely vacuolated cytoplasm of D. sylvatica and D. ranarum are the largely vacuolated trophozoite stages seen in D. kermiti n. sp. ( Fig. 1 B, D–E View Fig ; 2 B View Fig arrow). The third species, D. taiwanensis , described from the Alpine cricket frog Fejervarya limnocharis from Taiwan, is similar to D. kermiti n. sp. in that the trophozoite stages of both species contain vacuoles, the cytoplasm appears to be relatively coarse, and several measurements seem to overlap based on the scale provided. However, D. taiwanensis , similar to D. sylvatica , has meronts producing up to eight merozoites only. Likewise, the merozoites produced by D. taiwanensis are morphologically different from those of D. kermiti n. sp.: the nuclei seem to be larger and more prominent in comparison to the rest of the merozoites.
From the comparison of D. kermiti n. sp. to the currently recognised congeners infecting anurans, it is clear that these parasite species share close or overlapping morphology. Hence, since dactylosomatids possess several life stages in the vertebrate host, species can easily be misidentified. For example, gamont stages between the different species have largely comparable morphology and morphometrics. However, after careful review, screening a large number of infected hosts, comparing the stages observed in different individuals from different sites and seasons, we propose D. kermiti n. sp. as a new species.
| NMB |
Naturhistorishes Museum |
| MN |
Museu Nacional, Universidade Federal do Rio de Janeiro |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.