
Mesocapnia aptera, Lee & Baumann, 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.4760295 |
|
DOI |
https://doi.org/10.5281/zenodo.4763933 |
|
persistent identifier |
https://treatment.plazi.org/id/039F87F4-9976-FFF1-FC1F-F919FD834B91 |
|
treatment provided by |
Felipe |
|
scientific name |
Mesocapnia aptera |
| status |
sp. nov. |
Mesocapnia aptera View in CoL sp. n.
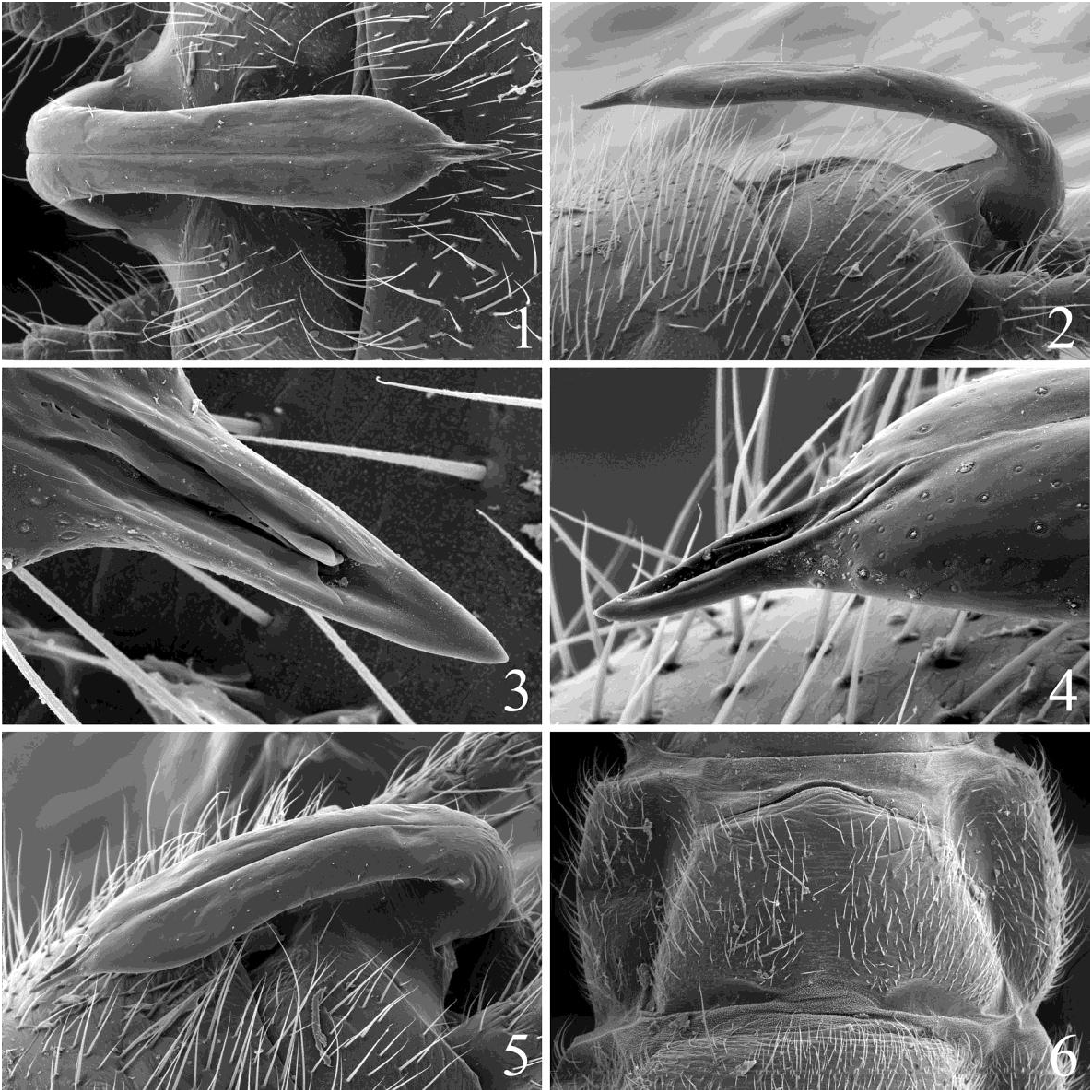
( Figs. 1-6 View Figs )
Material examined. Holotype ♂ and allotype ♀, USA, California, Mendocino Co., spring, upper Burger Creek , Dos Rios Road approximately 2.5 km east of Laytonville, 17-I-2011, J.J. Lee, deposited at the California Academy of Sciences, San Francisco, California . Paratypes: same data as holotype, 7♂, 17♀ ; same locality, 7-II-2011, J.J. Lee, 10♀ . Paratypes deposited at the Brigham Young University Collection and the Jonathan J. Lee Collection.
Male. Apterous, body length 4.0–5.0 mm. Body and appendages brown to dark brown, bearing numerous short setae. Antennae elongate, approximately same length as body. Pronotum with dark brown rugosities, meso-metanota with pair of shallow depressions centered in posterior quadrants of discs. Mesothoracic postfurcasternal plates separated from spinasternum. Abdominal terga with narrow, anterior dark brown band, segments 7-9 bearing stronger setae than anterior segments; segments 1-9 unmodified, tergum 10 cleft. Epiproct reflexed, extending to middle of segment 9 ( Figs. 1, 2, 5 View Figs ); in dorsal aspect epiproct broad ( Fig. 1 View Figs ), nearly parallel sided but widest at 3/4 distance from base to apex, length about 4 times greatest width; posterior portion slightly narrowed from broad base, gradually widening to anterior 1/4 then tapering to acute, apicomedian spine; spine with dorsomedial groove evident in SEM micrographs ( Figs. 3, 4 View Figs ). In lateral aspect ( Fig. 2 View Figs ) epiproct narrow, slightly convex dorsally, straight ventrally, apical spine in line ventrally. Cerci elongate, approaching abdomen length, at least 13 segments, segments bearing short setae and one stronger distoventral seta.
Female. Apterous, body length 5.0–7.0 mm. Body coloration and external morphology similar to male but lacking dark abdominal segment banding and stronger setae on segments 7-9. Abdominal terga fully sclerotized. Posterior margin of sternum 8 recessed medially to pleural folds ( Fig. 6 View Figs ), posterior border of subgenital plate ranging from slight median process to distinct blunt median point ( Fig. 6 View Figs ); anteromedian recessed area present; distinct thin, pale band usually evident from anterior recessed area to posteromedian process, pale band lacking setae as shown by SEM micrograph ( Fig. 6 View Figs ). The band is always present, represented by an absence of setae, even though not pale in every case.
Larva. Unknown.
Etymology. The specific epithet emphasizes the complete absence of wings in both sexes.
Diagnosis. Mesocapnia aptera superficially resembles the western North American genus Paracapnia based on its small size, coloration, and lack of wings. Males can be easily distinguished from Paracapnia by the apex of the epiproct that bears a small, stout spine typical of the genus Mesocapnia . The apterous condition will separate M. aptera from all other described Mesocapnia south of Alaska. We were unable to examine M. bergi for this study, however, the second author previously examined the types of M. bergi ( Baumann and Gaufin 1970) . Details of the epiproct shape will separate M. aptera from M. bergi . The epiproct of M. aptera is broader in dorsal aspect and narrower in lateral aspect than the epiproct of M. bergi . We examined the epiprocts of four additional Mesocapnia species using SEM micrographs and calculated the length to greatest width ratios. The species examined were M. autumna ( Baumann and Gaufin 1970) , M. frisoni ( Baumann and Gaufin 1970) , M. porrecta ( Jewett 1954a) , and M. yoloensis ( Baumann and Gaufin 1970) . The epiproct of M. frisoni was most similar to M. aptera in shape and ratio with a length to width ratio of 6.4 compared to 4.0 for M. aptera . Female M. aptera can be distinguished from other described western North American Capniidae , except Paracapnia baumanni Kondratieff and Lee 2010 , by the apterous condition and complete sclerotization of abdominal terga. Mesocapnia aptera and P. baumanni females strongly resemble each other but close examination of the subgenital plate will separate them. The subgenital plate of M. aptera has a posteromedian bluntly pointed process and a longitudinal, pale median stripe, while the subgenital plate of P. baumanni lacks these characters. For female M. aptera specimens lacking a well-defined subgenital plate process and obvious pale stripe, associated males may be needed for a positive determination.
Biological notes. Mesocapnia aptera specimens were collected along a spring creek flowing through a meadow at an elevation of ca. 730 meters. The spring creek channel, on 17 January 2011, was one meter wide with poorly defined, extended marshy margins. On 7 February 2011, after an extended dry period, water had receded from the marshy margins and was present only in the meter wide channel. The climate in the region is semi-Mediterranean. Normally, summers are warm and dry; winters are wet with most of the precipitation falling from November- April. We do not know if this spring creek is perennial. Stoneflies collected with M. aptera include Bolshecapnia maculata ( Jewett 1954b) , Capnia quadrituberosa Hitchcock 1958 , and an as yet undetermined member of the Capnia californica complex.
Remarks. Six additional apterous capniids are known from northern California, including: Paracapnia disala ( Jewett 1962) , Capnia lacustra Jewett 1965 (known only from Lake Tahoe and not examined for this study), Capnia fialai Nelson and Baumann 1990 , Paracapnia boris Stark and Baumann 2004 , Paracapnia humboldta Baumann and Lee 2007 , and Paracapnia baumanni . The males of these species are easily separated by epiproct shape. In females, the degree of abdominal tergal sclerotization may be of value as a taxonomic character. Jewett (1962) noted that the female of Paracapnia disala was apparently unique in the complete sclerotization of the first abdominal tergite. This character holds for known western apterous Paracapnia and for C. fialai . Additionally, P. humboldta females have complete sclerotization of the eighth abdominal tergite ( Baumann and Lee 2007), a character also noted by Hanson (1946) for some Allocapnia species found in eastern North America. Paracapnia baumanni females have all abdominal terga completely sclerotized ( Kondratieff and Lee 2010) as in Mesocapnia aptera . Complete abdominal tergal sclerotization also occurs in females of some apterous species of the Palearctic genus Capnioneura ( Vinçon and Sivec 2011; G. Vinçon personal communication).
A possible scenario leading to female tergal sclerotization involves habitat persistence (habitat stability over time), loss of flight capacity and wings, and the need for protection of the dorsal abdomen. Roff (1986) and Dudley (2000) suggest habitat persistence as a major factor underlying reduction of insect flight capacity. Brodsky (1994) suggests Plecoptera forewings provide abdominal protection at rest. The degree of female abdominal tergal sclerotization in apterous stoneflies may be evolving to protect the dorsal abdomen. Smith et al. (2001), discussing aquatic mite zoogeography in North America, found some “Tertiary relict distributions, suggesting that they originated in Laurasia”. Aquatic mite “Tertiary- relict” distributions include unglaciated refugia such as coastal California and Oregon. Perhaps the western apterous stonefly species are relictual populations, having persisted in refugia, allowing for loss of wings and increased abdominal tergal sclerotization.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.