
Parotoplana
|
publication ID |
https://doi.org/ 10.5281/zenodo.177648 |
|
DOI |
https://doi.org/10.5281/zenodo.6242272 |
|
persistent identifier |
https://treatment.plazi.org/id/039FC129-1E0C-FFE8-FF61-6ADE88B6FDB7 |
|
treatment provided by |
Plazi |
|
scientific name |
Parotoplana |
| status |
|
Parotoplana sp. B.
( Fig. 5 View FIGURE 5 C; Fig. 7 View FIGURE 7 E)
Material: one whole mount (CZM 58). Apulia, Italy: Porto Cesareo (Lecce) Bacino Grande bay, loc. Scala di Furno (lat. 40°16’26”N, long. 17°52’53.68”E), slightly silty coarse sand among rocks, lower intertidal in the Otoplana -zone. May 2005.
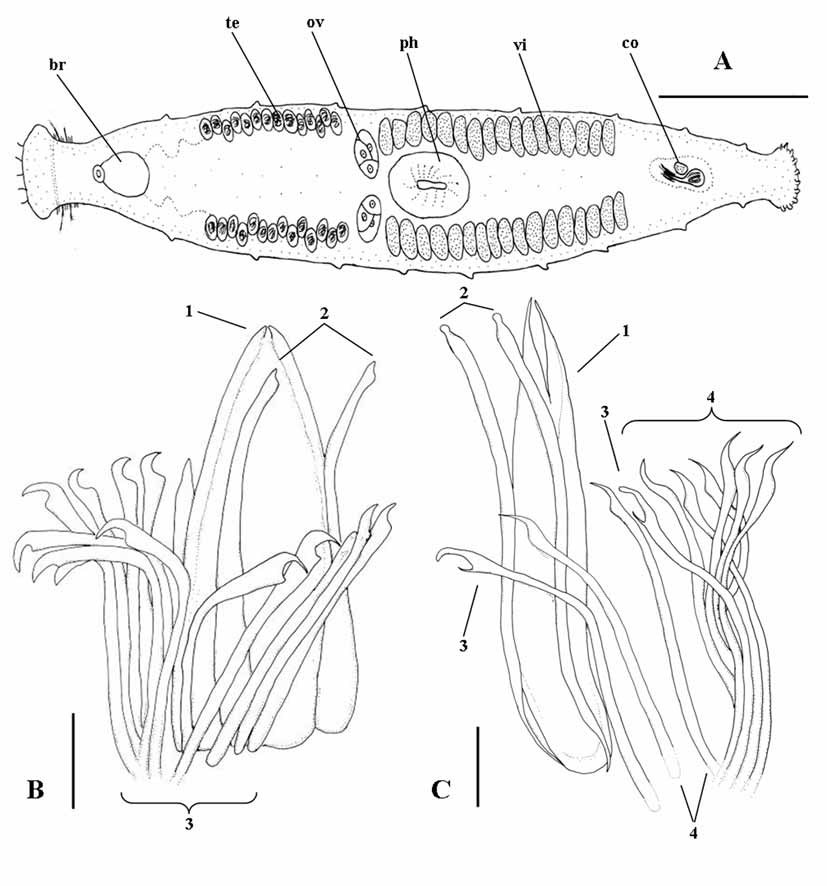
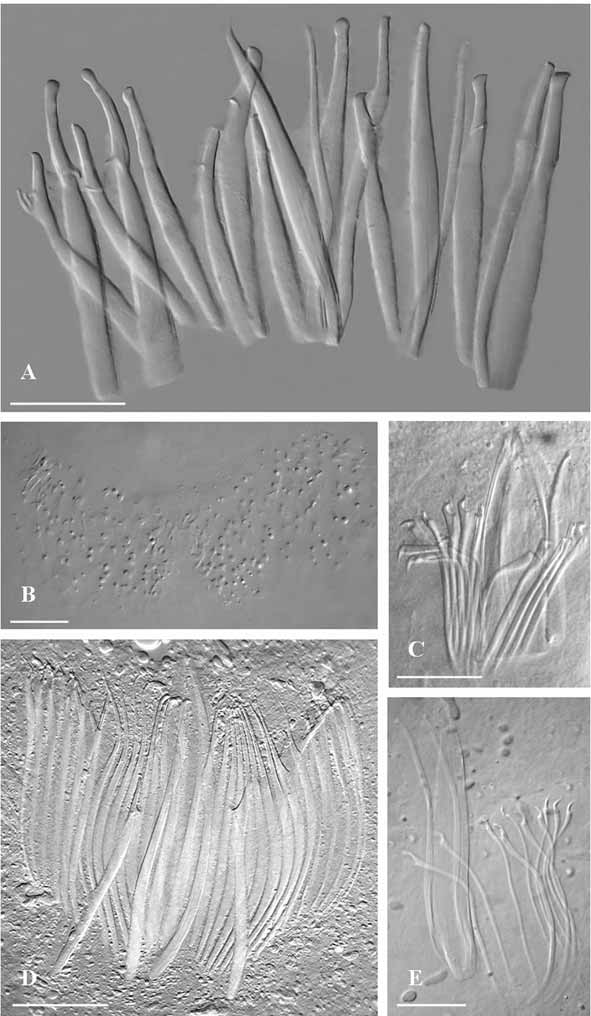
Description. General morphology of the body and arrangement of organs similar to the other species of the genus. Sclerotized apparatus ( Fig. 5 View FIGURE 5 C) consisting of:
1) a central plate-like stylet (115 ìm long) with distinctly forked apex;
2) two straight companion spines (105–106 ìm long), slightly swollen distally, and forming a markedly bulbous distal point;
3) two spines (82 µm) with sickle shaped apices, obtuse distal tips, and with subterminal, narrow and elongate teeth;
4) eight spines (75 to 78 µm long) with sickle shaped apices, pointed distal tip, and obtuse subterminal teeth.
Karyotype. Chromosome number: n = 6; FN = 12. All chromosomes are isobrachial; Chrom. VI is submetacentric with very low index. The first two pairs are distinctly larger than the remaining chromosomes. Chrom. I = r.l.: 32.23 + 2.85; c.i.: 46.97 + 0.24 (m); Chrom. II = r.l.: 27.46 + 2.95; c.i.: 38.46 + 3.08 (m); Chrom. III = r.l.: 12.82 + 0.38; c.i.: 41.93 + 0.39 (m); Chrom. IV = r.l.: 10.47 + 0.18; c.i.: 37.11 + 2.48 (sm); Chrom. V = r.l.: 8.75 + 1.05; c.i.: 44.49 + 3.22 (m); Chrom. VI = r.l.: 8.23 + 0.38; c.i.: 26.93 + 1.69 (sm) (based on the measurements of three spermatogonial plates).
Remarks. The two specimens, although similar in the general arrangement of the sclerotized structures and karyotype, do not appear to be conspecific. In fact, they differ in the number of spines (14 in Parotoplana sp. A, 12 in Parotoplana sp. B) and for their size (consistently larger in Parotoplana sp. B). Furthermore, their stylet is different (plate–like in Parotoplana sp. A; distally split in Parotoplana sp. B), as is the shape of the companion spines, forming a unique bulbous apex in Parotoplana sp. B. This latter specimen also showed a peculiar morphology of the first, larger pair of spines of the girdle, namely an exceptionally long and thin subterminal tooth.
Among the species found in southern Apulia, the two specimens are similar to P. pythagorae sp. n. in number and arrangement of spines. This species however has a forked stylet, patently formed by two spines joined by a thin membrane; the companion spines are distinctly broader than the others and provided with characteristic apical triangles, and the girdle, though similar to Parotoplana sp. A, is formed by larger spines. The comparatively large sample of P. pythagorae sp. n. examined showed constant characters, and was syntopic with Parotoplana sp. B.
The two specimens appear to be very similar to two Mediterranean species, P. macrostyla from the Ligurian Sea and P. renatae from the Gulf of Lyon, because of the presence of a median, laminar stylet, two larger companion spines, and a girdle of 10–13 spines. According to Lanfranchi (1978), the discriminating feature between P. re n a t a e and P. macrostyla is the absence of a common female duct in the latter. In P. macrostyla , the oviducts join just before their opening into the common genital atrium – a situation similar to P. spathifera sp. n. described above. However, it must be noted that the distal joining of the oviducts is impossible to appreciate on semi–squashed living specimens, and easily overlooked in less than very high quality sections. P. renatae , on the other hand, is described as provided with a triangular stylet, whereas P. macrostyla has a distally forked stylet. However, the shape of the stylet can be unequivocally assessed only in strongly squeezed mounts. Based on available data, one might therefore question whether P. renatae and P. m a c ro s t y l a are indeed two different species. The problem will be only solved following a detailed study of further specimens from the type localities.
The two specimens from Porto Cesareo seem to add further complexity to the question. In fact, at least Parotoplana sp. B, though similar to P. m a c ro s t y l a for the shape of stylet, differs in the unique morphology of companion spines and the two largest spines of the girdle. Parotoplana sp. A has a triangular plate-like stylet, similar to that described for P. renatae , but differs for the less acutely pointed companion spines and the hooked shape of most of the girdle spines. Both specimens show a karyotype indistinguishable to that of P. macrostyla (cf Curini-Galletti et al., 1984) for the general morphology of chromosomes, and for the shape of the smallest pair (Chrom. VI), which is distinctly less isobrachial than the others (ranging from submetacentric with very low index to subtelocentric).
The complexity of the taxonomy of the group, the paucity of the material available for study, and the lack of histological sections, refrain us from any further taxonomic inference on the two specimens.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.