
Tanypus formosanus sensu Niitsuma (2001)
|
publication ID |
https://doi.org/ 10.5281/zenodo.198648 |
|
DOI |
https://doi.org/10.5281/zenodo.5625464 |
|
persistent identifier |
https://treatment.plazi.org/id/03A08A1F-FF90-FFA9-FF7F-FB17B6980935 |
|
treatment provided by |
Plazi |
|
scientific name |
Tanypus formosanus sensu Niitsuma (2001) |
| status |
|
Tanypus formosanus sensu Niitsuma (2001) View in CoL
Niitsuma (2001) stated that the species reported or described previously in Japan as T. punctipennis Meigen, 1818 is T. formosanus ( Kieffer, 1912a) . This thesis is herein revised.
The descriptions on T. formosanus by Kieffer (1912a; 1916a, b) were based on adults only, and there had been no descriptions of immature stages published before Niitsuma’s report (l.c.). Thus, identification of immatures based on the early descriptions is impossible, and the differences in adult characteristics of T. formosanus based on Kieffer’s descriptions and the material examined are uncertain. The evidences by Niitsuma (2001) are stated in the ‘Introduction’ of his article. Two characters are pointed out for separation the two species compared - the length of palpomeres and empodium. According to Niitsuma (l.c.) ‘they (his specimens) differ from T. punctipennis in the palpal segment III, which is longer than segment II, and in the empodium, which is half as long as the claws. In the latter species ( T. punctipennis ), palpomeres II and III are subequal ( Sublette 1964) and the empodium is as long as the claws ( Roback 1966). The features of the palp and empodium match those of T. formosanus described by Kieffer, better than they do those of T. punctipennis . ’ First, the Sublette’s report (1964) does not include data on T. punctipennis , but the two new species, i.e. T. neopunctipennis and T. carinatus based on the specimens from the United States, which had been misidentified with T. punctipennis by Malloch. Therefore, Niitsuma’s quotation about the lengths of palpomere is ineffective. Secondly, concerning the empodium character, Kieffer’s description of T. formosanus expresses ‘claws twice as long as empodium’, and Roback 1966 described ‘empodium about as long as claws’ based on the type of T. punctipennis from Europe. However, empodium is a fine hair-like structure, indistinct even under high magnification. It is questionable, whether length of empodium is a stable character, or whether the length of this structure could be the only character for separation the species. Presumably, Niitsuma (2001) did not know the key character of wing patterns advocated by Langton and Pinder (2007).
Niitsuma (2001) also evidenced that his adult specimens belong to T. formosanus , not to T. kraatzi . He listed up three characters: in T. kraatzi ‘(i) the adult with a relatively high foreleg ratio (higher than 1), (ii) a bare, slender gonostylus in the male hypopygium ( Kieffer 1913, 1918; Goetghebuer 1936), and (iii) the larva with six to seven lateral teeth on the dorsomentum ( Zavřel and Thienemann 1921, Fittkau and Roback 1983)’. Regarding (i), ‘fore tibia equal ta1’ by Kieffer (1913: 15); ‘fore tibia not longer than the ta1’ by Kieffer (1918: 165), and by Goetghebuer (1936: 7). However, the European specimen of T. kraatzi ( Germany, Plön) shows 0.87 of fore leg ratio. This ratio varies between 0.79–0.90, 0.86 (16) in spcimens of T. kraatzi presently examined ( Table 4). In the other specimens having measurable foreleg, identified as T. kraatzi by Fittkau or Reiss, the ta1 is always shorter than the tibia (Spies, pers. comm.). Concerning (ii), the specimen of ZSM mentioned above, has 14 setae at least on the gonostylus. Similarly, Langton and Pinder (2007, vol. 2: fig. 129 C) illustrated gonostylus of T. kraatzi bearing setae. Gonostylus of T. kraatzi from Russia is also never bare (Sergeeva, pers. comm.). According to Spies (pers. comm.), Kieffer had often received the specimens from Thienemann in alcohol vials, thus, one possibility is that the longer setae on the gonostyli were rubbed off, so that the gonostylus appeared ‘bare’ under low magnification. With respect to (iii), number of dorsomental teeth is a variable character. According to Vallenduuk and Pillot (2007), there are ‘usually 6 dorsomental teeth (rarely up to 8)’ in T. kraatzi . In this study, 2/12 of the specimens of T. kraatzi examined have 5 teeth, whereas the remaining have 6 teeth. Measurements of ligula and paraligula of the specimen of T. kraatzi from Germany ( ZSM) are as follow: length of ligula 80μm, basal width 39μm, minimum width 30μm, ration of length to basal width 2.05, paraligula with 2 branches, dorsomentum with 7 teeth on each side. These data agree with those of T. kraatzi by the present author. After all, the evidences by Niitsuma (2001) in order to separate T. kraatzi and T. formosanus should be regarded as unreliable. Regarding characters of larval dorsomental teeth, ligula, paraligula, and other structures, the differences also could not be recognized between Niitsuma’s specimens and the descriptions of T. kraatzi by Zavřel and Thienemann (1921), Fittkau and Roback (1983) and Makarchenko (2006). Taken together, no contradiction was found between the results by the present author and the previous descriptions or keys to T. kraatzi in all stages.
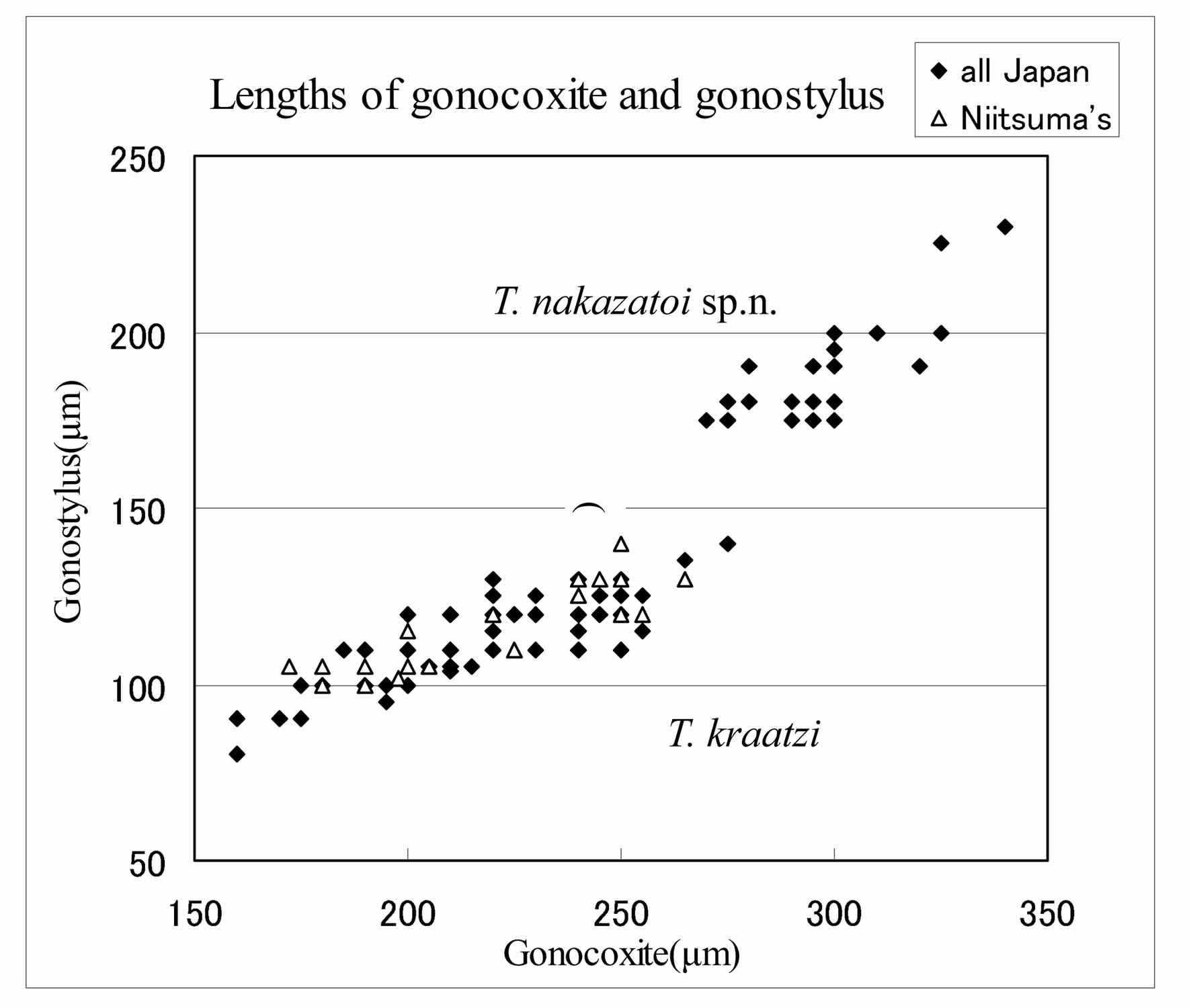
It is easy to identify the specimens accurately. All morphological data of Niitsuma’s specimens ascribed to T. formosanus show coincidence with those of T. kraatzi (e.g. Fig. 35 View FIGURE 35 ). T. punctipennis by Tokunaga (1937a) also appears to belong to T. kraatzi , because of the wing spots distance (Plate 2, Fig. 19 View FIGURES 19 – 24 Tokunaga 1937a), HR 2.0 (based on Plate 2, Fig. 28 View FIGURES 25 – 29 ) and total length of male (1.8–3.3mm).


| ZSM |
Bavarian State Collection of Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |