
Psilotris, GINSBURG, 1953
|
publication ID |
https://doi.org/10.1111/zoj.12394 |
|
publication LSID |
lsid:zoobank.org:pub:E952647E-1571-4A14-8BD4-54D1746760D0 |
|
persistent identifier |
https://treatment.plazi.org/id/03A0C25D-BB73-FF8C-B7A8-FCEFFAC1EC1B |
|
treatment provided by |
Marcus |
|
scientific name |
Psilotris |
| status |
|
PSILOTRIS GINSBURG, 1953 View in CoL View at ENA
TYPE SPECIES: PSILOTRIS ALEPIS GINSBURG, 1953 ( BY ORIGINAL DESIGNATION)
Diagnosis
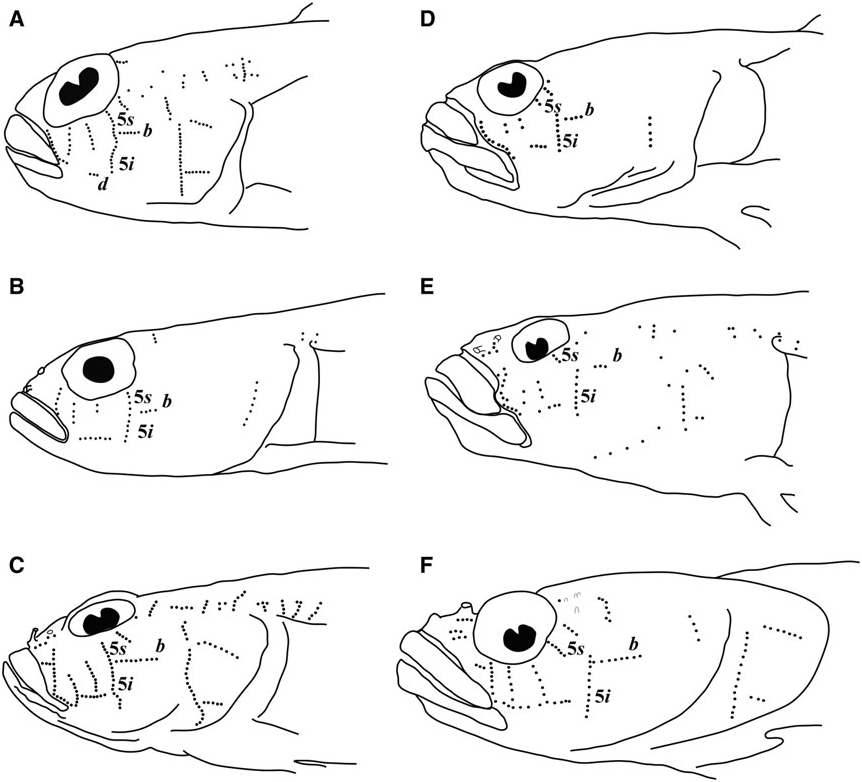
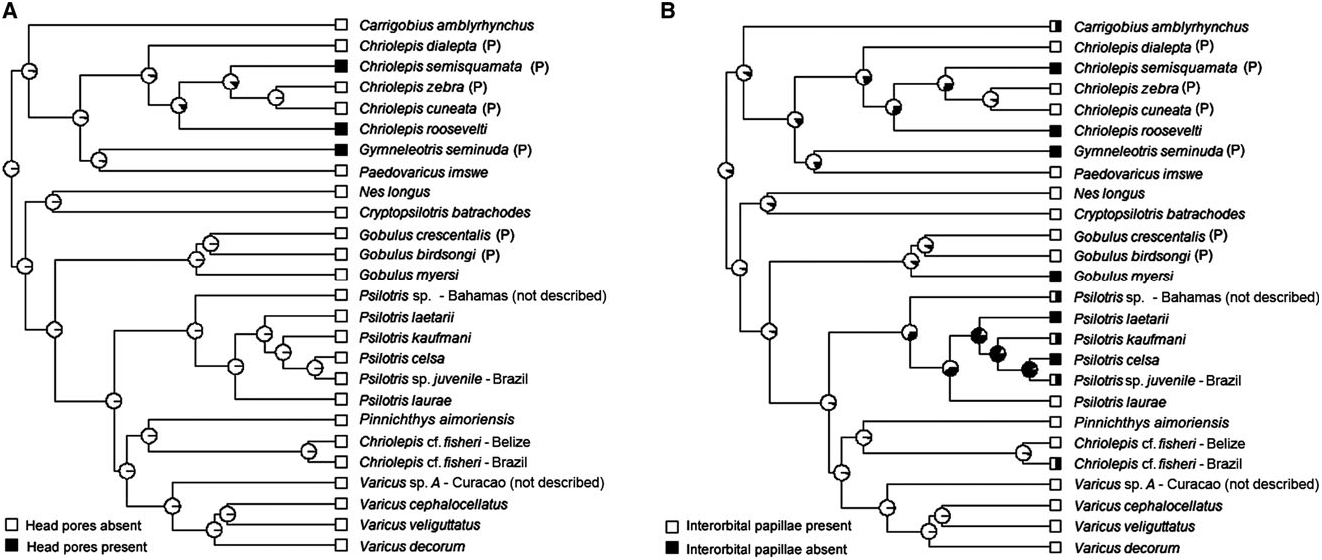
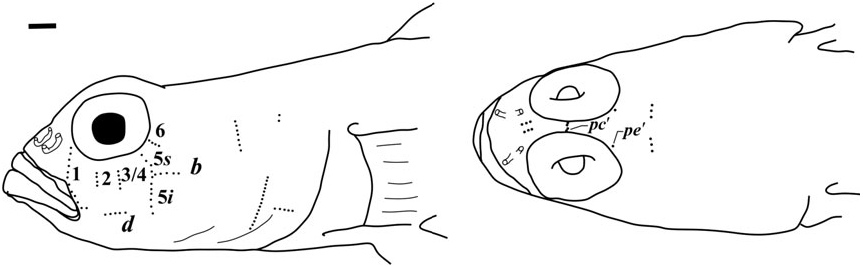
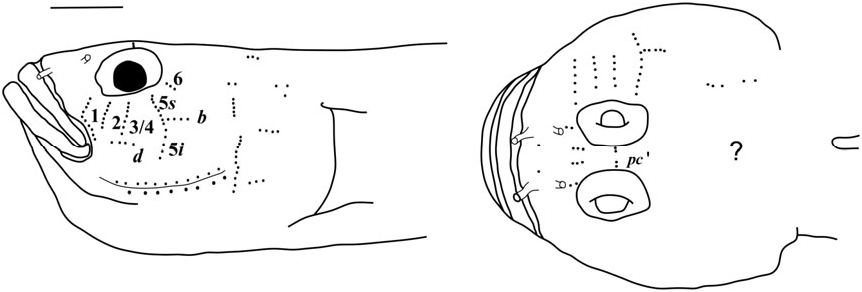
Possesses all taxonomic characters present in most members of Gobiosomatini and the Gobiosoma group Sensory papillae ( Fig. 16 View Figure 16 ): A transverse pattern with rows 1, 2, 3/ 4, 5s /5i, and 6 present; rows 2 and 3/4 short; row 5s/5i joined forming a single row and extending below the level of row d; row b short with five or six papillae, ending anteriorly at row 5s/5i; interorbital row pc’ with two papillae, row pe’ with a single papilla.
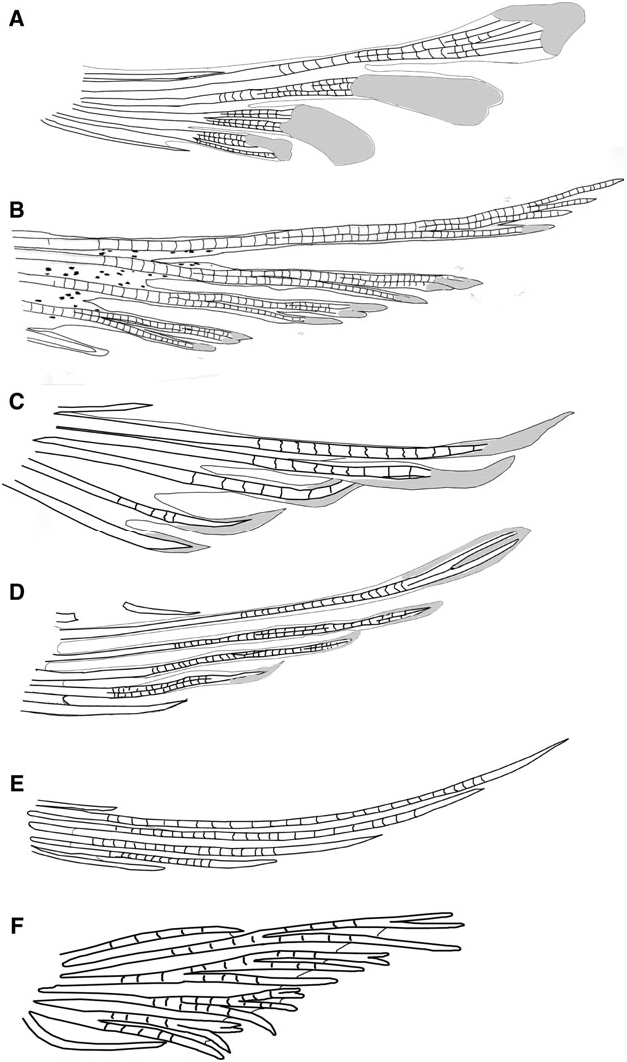
Vertebral skeleton: Dorsal pterygiophore formula 3 – 221110; two anal-fin pterygiophores inserted anterior to first haemal arch; hypurals 1 and 2 fused to hypurals 3 and 4 along one-quarter of length; 27 vertebrae – 11 precaudal and 16 caudal.
Habitat: Holotype was collected on a calcareous sand and dead Halimeda spp. algal rubble zone, with scattered shallow calcareous rock ledges.
Distribution: Known only from San Salvador, Bahamas, the type locality.
Etymology: The name saurimimica is derived from ‘ saurus ’, a genus of lizard fish and the Greek ‘ mimic ’, as the colour pattern closely resembles that of a lizard fish.
(first dorsal-fin spines VII, pterygiophore insertion pattern of 3 – 221110; 27 vertebrae – 11 precaudal and 16 caudal; hypurals 1 and 2 fused to some extent with hypurals 3 and 4 and the terminal vertebral element, one epural); pelvic fins well separated, lacking both anterior frenum and well-developed membrane connecting innermost rays; pelvic-fin rays 1 – 4 branched, typically without fleshy or flattened tips (sometimes slightly fleshy in Psilotris boehlkei Greenfield, 1993 ), fifth pelvic-fin ray unbranched; pelvic-fin rays extending posteriorly from about halfway to anus to anus, but never beyond anus; body lacking scales (modified basicaudal scales present in one undescribed species; see Remarks below); two anal-fin pterygiophores inserted before first haemal spine in all but one species ( Psilotris laurae sp. nov., Van Tassell, Tornabene & Baldwin, 2016), which has one anal-fin pterygiophore inserted before first haemal spine; papillae rows 5i and 5s connected in some species, not connected in others; cephalic lateralis canals and pores absent; second dorsal-fin rays I,9 – 10; anal-fin rays I,7 – 10; body coloration highly variable (presumably for camouflage with a specific substrate type), but frequently with vertical bars (broken or continuous, yellow to dark brown or black) or saddles over body and head, some species with prominent spots or mottling over side of body. All six valid species occur from the western Atlantic Ocean.
Remarks
Psilotris View in CoL was described by Ginsburg (1953) for Psilotris alepis View in CoL . Like Chriolepis View in CoL , Psilotris View in CoL was considered by Ginsburg as belonging to the Eleotridae View in CoL for the split pelvic fins lacking a connective membrane and anterior frenum, a character now known to have evolved multiple times in Gobiidae View in CoL . In addition to the split pelvic fins, Psilotris View in CoL lacks cephalic lateralis pores and body scales – two characters that appear multiple times across the Gobiosomatini . The scaleless and pore-less Psilotris celsa Bohlke, 1963 View in CoL and Psilotris batrachoides View in CoL were added to the genus, and Bohlke noted that the three species were ‘more different in appearance than members of most shorefish genera’ (Bohlke, 1963: 2). Later, Psilotris View in CoL kauf- mani Greenfield, Findley & Johnson, 1993; Psilotris bohlkei Greenfield, 1993 ; and Psilotris amblyrhynchus Smith & Baldwin, 1999 View in CoL were also added to the genus.
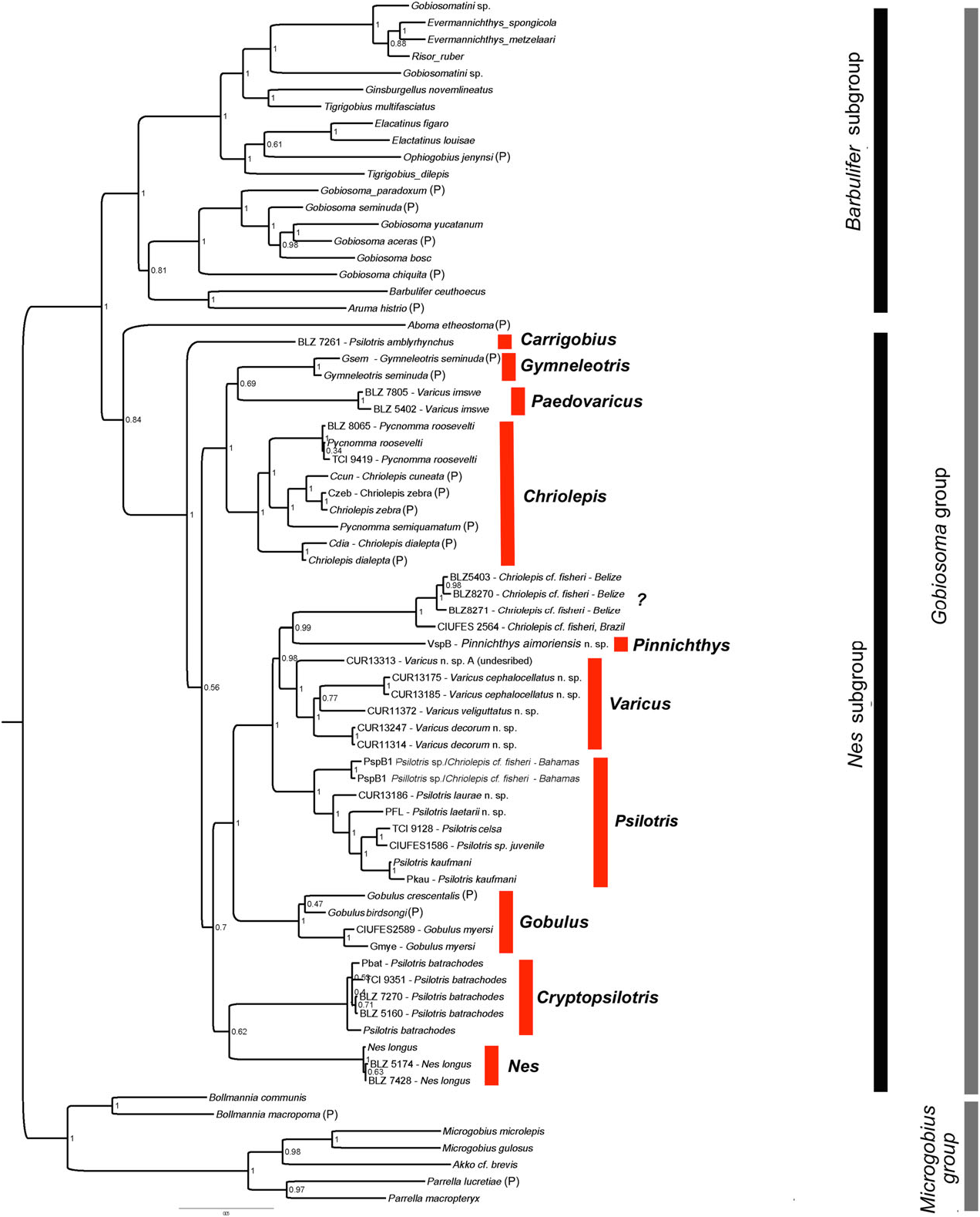
Smith & Baldwin (1999) and Greenfield (1993) recognized that the species of Psilotris View in CoL probably do not form a monophyletic group. In addition to not being united by unique shared derived characters, Psilotris batrachodes View in CoL has a colour pattern and general appearance that is strikingly different from the other members of the genus, and Psilotris amblyrhynchus View in CoL has a notably different body shape, with a blunt anterior profile versus a gradually sloped or pointed anterior profile. In our molecular phylogeny ( Fig. 5 View Figure 5 ), Psilotris amblyrhynchus View in CoL and Psilotris batrachodes View in CoL are recovered well outside the clade containing Psilotris celsa View in CoL , Psilotris kaufmani View in CoL , and several other undescribed species. The new genera Carrigobius View in CoL and Cryptopsilotris View in CoL are erected for Psilotris amblyrhynchus View in CoL and Psilotris batrachodes View in CoL , respectively (see remarks section of each genus for additional information). The steep anterior profile and branched fifth pelvic-fin ray (in adults) distinguish Carrigobius View in CoL from Psilotris View in CoL , and the body coloration and low anal-fin ray counts (I,6 – 7) distinguish Cryptopsilotris View in CoL from Psilotris View in CoL .
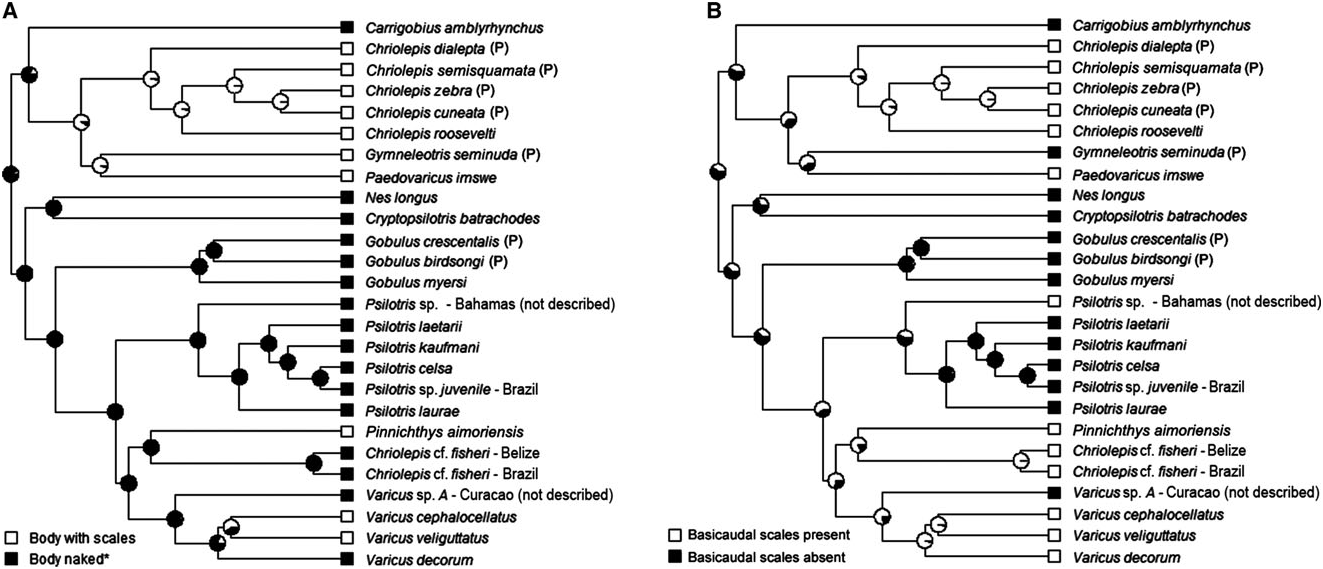
The loss of body scales and cephalic lateralis pores have apparently occurred multiple times within the Nes subgroup ( Figs 7A View Figure 7 and 9A View Figure 9 ). In addition to Psilotris , Carrigobius , and Cryptopsilotris possessing these losses, at least two species in the genus Varicus are also naked and without pores ( Varicus decorum sp. nov. and an undescribed species). Because of the parallel loss of scales (or conversely, the regaining of scales in some members of Varicus ), we know of no characters that unambiguously distinguish Varicus from Psilotris . Although there are general trends in several characters, there are exceptions for all of them. For example, of the ten species of Varicus recognized in this study (nine named herein plus one undescribed), eight possess scales on the body, whereas all species of Psilotris are naked. Of the six species of Psilotris considered valid in this study, all but one, Psilotris laurae sp. nov., have two anal-fin pterygiophores inserted before the haemal arch, whereas all Varicus have one. All species of Psilotris have pelvic-fin rays 1 – 4 branched without expanded fleshy tips, whereas only two of the ten known species of Varicus [ Varicus benthonis ( Ginsburg, 1953) and Varicus vespa Hastings & Bortone, 1981 ] have pelvic-fin rays that are branched to the tips (although these two have very small fleshy pads at the tips of some branches). Lastly, all ten of the known species of Varicus occur at or below 60 m, whereas only one species of Psilotris ( Psilotris laurae sp. nov.) occurs on deep reefs. Despite partial overlap in all morphological characters examined here, the two genera form distinct clades in our molecular phylogeny with strong support (Bayesian posterior probability = 1.0).
In addition to the two new species of Psilotris described here ( Psilotris laurae sp. nov. and Psilotris laetarii sp. nov.), there are two additional lineages in the Psilotris clade that are of uncertain identity. One juvenile specimen of Psilotris from Brazil is recovered as sister to Psilotris celsa . It is not clear whether this specimen is truly Psilotris celsa , which would expand the range of the species considerably, or if it represents an undescribed species. Another lineage comprises two specimens from the Bahamas, and could represent an undescribed species. This latter species differs from all other Psilotris in possessing two modified basicaudal scales, which makes it superficially resemble the incertae sedis species Chriolepis fisheri . The second dorsal-fin ray and anal-fin ray counts (I,9 and I,8, respectively) of this species do not match that of the Chriolepis fisheri holotype (I,10 and I,9). The Psilotris specimens from the Bahamas may be the same species as the ‘ Chriolepis cf. fisheri ’ described by Smith-Vaniz & Jelks (2014). These specimens from the Bahamas also superficially resemble another species in our phylogeny that is recovered as sister to Pinnichthys . This latter species also possesses only basicaudal scales and has second dorsal-fin and analfin ray counts more closely matching those of the Chriolepis fisheri holotype; however, additional specimens and genetic data are needed to resolve the status of Chriolepis fisheri and the putatively new species of Psilotris from the Bahamas (see the remarks section for Chriolepis above).
PSILOTRIS LAETARII VAN TASSELL & YOUNG SP. NOV. BURROW SPLITFIN GOBY
FIGS 17 – 19 View Figure 17 View Figure 18 View Figure 19
Holotype
AMNH 261272 About AMNH , 23.5 mm SL, female, Marathon , Florida, USA, 24.39.470N – 81.00.397W, 7.6 m depth, 20 September 2012, John ‘Bucky’ Wile III.
Paratypes
AMNH 264217 About AMNH , 21.5 mm SL, male, collected with holotype ; AMNH 257973 About AMNH , 23.27 mm SL, male, Marathon , Florida USA, 24.39.470N – 81.00.397W, 7.9 m depth, 10 September 2014, Warren ‘Skip’ Wohlers ; AMNH 257974 About AMNH , 19.3 mm SL, male, Marathon , Florida USA, 24.39.470N – 81.00.397W, 7.9 m depth, 15 June 2013, John ‘Bucky’ Wile III .
Diagnosis
Side of body and head without scales; no modified basicaudal scales present; first dorsal fin VII, without elongate spines; second dorsal I,9 – 10; anal fin I,7 – 8; pelvic fins well separated, anterior frenum absent, membrane connecting fifth pelvic-fin rays absent or very low; fifth pelvic-fin ray unbranched and half the length of fourth; pelvic-fin rays without fleshy tips; papillae row 5s and 5i connected forming a single row, interorbital papillae row pc’ present; head and preopercle canals and pores absent; two anal-fin pterygiophores inserted anterior to haemal arch.
Description
Morphometric data are presented in Table 3.
Median and paired fins: First dorsal fin VII(4), spines 1 – 3 about equal in length, spines 4 – 6 slightly shorter, spine 7 three-quarters the length of first spine; second dorsal fin I,9*(1), I,10(3); anal fin I,7* (1), I,8(3); pectoral-fin rays 15(1), 16(2), 17*(1), elongate, extending to a vertical through second dorsal-fin ray 1 or 2; pelvic fin I,5(4); pelvic fins well separated, lacking anterior frenum, membrane connecting fifth rays absent or very low; fourth pelvic-fin ray longest, extending three-quarters distance to anus; rays 1 – 4 branched two or three times, fifth ray unbranched and half the length of fourth ray; tips of pelvic-fin rays not ending in fleshy pads; caudal fin ovate; branched caudal-fin rays 15 (3); segmented caudal-fin rays 17(4).
Scales: Scales on head and trunk absent.
Head: Jaw extending posteriorly to a vertical through the anterior margin of eye; anterior nare a short tube, posterior nare an opening with raised rim; no cephalic lateralis pores on head or preopercle; eyes 24.1 – 29.1% HL, dorsolateral; interorbital narrow, 10.7 – 14.4% HL; upper jaw with two or three rows of teeth at anterior, outer row extending to posterior end of premaxilla, teeth large but slightly smaller than teeth in lower jaw, conical with recurved tips; lower jaw with two or three rows at anterior, slightly larger than upper teeth, pointed conical and slightly recurved, inner row with four larger teeth at mid-dentary.
Genitalia: Male urogenital papilla conical and pointed; female urogenital papilla short, round, bulbous with small villi around edge.
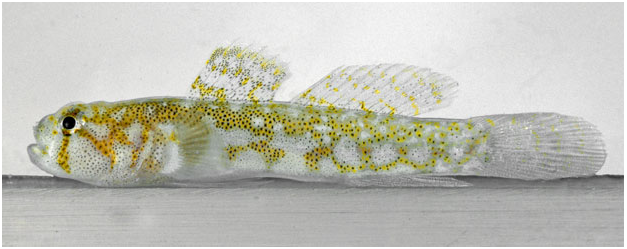
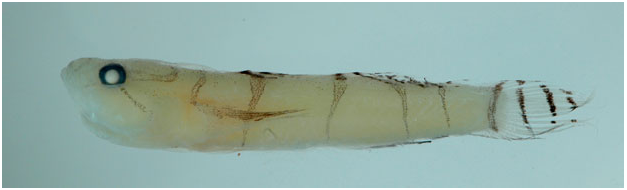
Colour in life ( Fig. 17 View Figure 17 ): Background colour of body and head whitish; head with three dark yellowish – orange bars, a vertical bar under eye extending to jaw, an oblique bar extending from eye to lower edge of preopercle, a third bar extending from rear of the eye to top corner of opercle; top of head with reticulating yellow – orange lines; trunk with a reticulum of dark yellow – orange lines, darker at the anterior, becoming lighter posteriorly; numerous scattered small dark concentrations of melanophores on head and trunk, more numerous on nape and on dorsal trunk under first dorsal fin, the concentrations of melanophores superimposed on the yellow – orange reticulum are larger and dark; first dorsal fin with three irregular, transverse bands of yellow spots, one at base of fin ending between fifth and sixth spine, a central band along middle of fin, upper band not extending to upper margin of fin; distal margin of fin clear; entire first dorsal fin with scattered melanophores, particularly concentrated on and adjacent to the yellow bands; second dorsal fin clear with five thin oblique light-yellow bands with scattered small melanophores, bands equally spaced along dorsal extent of fin but not extending to dorsal margin, which lacks pigment; anal fin unpigmented with a few widely scattered small melanophores; caudal fin translucent with five thin, irregular pale yellow bars; pectoral fin translucent, base of fin whitish with scattered melanophores and a yellow – brown spot at dorsal corner of base; pelvic fin lacking pigment.
Colour in preservation ( Fig. 18 View Figure 18 ): Background colour of head and body yellowish pale; all areas with yellowish – orange colour before preservation, now light brown or grey, and peppered with dark melanophores, including stripes on head, and reticulations on nape and trunk; first dorsal fin peppered with melanophores, loosely concentrated to form two lateral stripes, stripes less apparent posteriorly; second dorsal fin with very small and sparsely spaced irregular patches of melanophores; caudal fin immaculate; anal fin with very light dusting of small melanophores; pelvic fin immaculate.
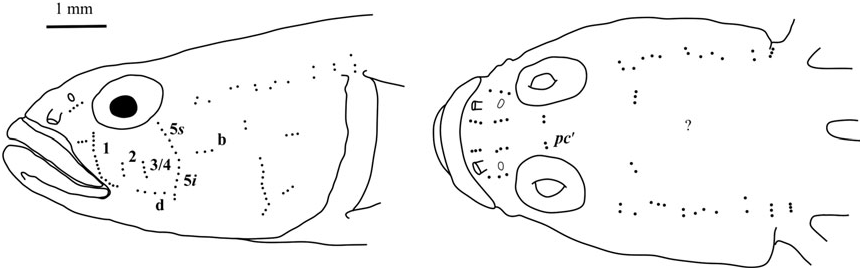
Sensory papillae ( Fig. 19 View Figure 19 ): Sensory papillae in a transverse pattern; transverse row 1 long, extending from orbit to row d; rows 2 and 3/4 short, extending from mid-cheek to near row d; rows 5s and 5i connected, extending from orbit to below level of row d; row b short, anterior end not reaching row 5s/ 5i; interorbital row pc’ with two papillae present.
Vertebral skeleton: Dorsal pterygiophores formula 3 – 221110; two anal-fin pterygiophores inserted anterior to first haemal arch; hypurals 1 and 2 fused to hypurals 3 and 4 along approximately one-half of their length; 27 vertebrae – 13 precaudal and 14 caudal.
Habitat: Types were collected from a sand and rubble substrate, rubble about 0.5 – 10.0 cm in diameter, located 45 – 90 m from the gradual reef drop-off. Found in or near burrows of Opistognathus aurifrons ( Jordan & Thompson, 1905) (yellowhead jawfish) at depths of 8.5 m.
Types were collected solely during dedicated collections of Opistognathus aurifrons (yellowhead jawfish) by use of a 1% quinaldine solution (2 – methylquinoline) at depths of 6.0 – 8.5 m. The only observance and collection of this species has been from within the yellowhead jawfish burrows. There appears to be some relationship between the two species, as this new goby is found in no other situation that the authors are aware of, but further research will be necessary to establish what that relationship may involve. The authors have not seen Psilotris laetarii sp. nov. exit from any other kind of burrow; such burrows are common among the back-reef rubble habitat. This is not an abundant species but can be found on a regular basis from April to November, when the jawfish are seasonally abundant.
Distribution: Known only from the type series collected off the south shore of Marathon, Florida.
Etymology: Named in honor of Heath Jens Laetari, 28 years old, Vice President of Dive Operations, Partner & Acquisition Manager for Dynasty Marine, who was lost at sea on 14 September 2006, doing what he loved to do.
Comparisons: Psilotris laetarii sp. nov. is the only known Psilotris species to have a reticulating yellowish orange pattern on the body. Psilotris celsa has vertical yellowish orange bars on the side of the body ( Fig. S4C View Figure 4 ), similar in colour to that of the reticulations on Psilotris laetarii sp. nov.; however, the shape of the overall pattern is different. In addition to the colour pattern, Psilotris laetarii sp. nov. differs from Psilotris alepis and Psilotris boehlkei in having papillae rows 5i and 5s connected (separate in Psilotris alepis and Psilotris boehlkei ). This species can be further distinguished from Psilotris boehlkei in having fewer anal-fin rays (I,7 – 8 vs. I, 9 in Psilotris boehlkei ), and in having a jaw that does not extend posteriorly past a vertical through the pupil. Both Psilotris laurae sp. nov. and Psilotris kaufmani have bicoloured pectoral fins, whereas the pectoral fin in Psilotris laetarii sp. nov. is uniformly unpigmented. Psilotris laetarii sp. nov. is distinguished from the scaleless Varicus species Varicus decorum sp. nov. both in colour pattern (yellow – orange reticulations versus round yellow spots in Varicus decorum sp. nov.) and in having pelvic-fin rays 1 – 4 that branch to the tips without fleshy pads (tips either unbranched or branched and re-fused, with fleshy tips in Varicus decorum sp. nov.).
PSILOTRIS LAURAE VAN TASSELL, TORNABENE & BALDWIN SP. NOV. THIN- BARRED GOBY
FIGS 20 – 22 View Figure 20 View Figure 21 View Figure 22
Holotype
USNM 426779 About USNM , 26.8 mm SL, male, off Bonaire City Dock , Kralendijk, Bonaire 12.15N – 68.2829W, 30 May 2013, 114 – 137 m, B. Van Bebber, A. Schrier, C.C. Baldwin & T. Christiaan. GoogleMaps
Diagnosis
No scales on sides of body or head; no modified basicaudal scales; first dorsal fin VII, with spines 1 – 4 slightly elongate; second dorsal fin I,9; anal fin I,8; elongate pointed pectoral fin with 18 rays; pelvic fins separate, no anterior frenum or membrane connecting fifth rays, rays 1 – 4 branched once, fifth ray unbranched and one-quarter the length of fourth ray; papillae row 5s/5i connected forming a single row at the anterior of row b, interorbital papillae row pc’ with two papillae; head and preopercle canals and pore absent; one anal-fin pterygiophore inserted anterior to haemal arch; a distinct colour pattern with five conspicuous thin bars along trunk, bars bright yellow in life or black in preserved specimens.
Description
Morphometric data are presented in Table 3.
Median and paired fins: First dorsal fin VII, spines 1 – 4 slightly elongate, second spine longest, tips of spines projecting from membrane; second dorsal fin I,9; anal fin I,8; pectoral fin 18, elongate, pointed, extending to second or third ray of anal fin; pelvic fins I,5; pelvic fins well separated, lacking both anterior frenum and membrane connecting bases of innermost pelvic-fin rays; fourth pelvic-fin ray longest, extending posteriorly to anus; rays 1 – 4 branched once, fifth ray unbranched and onequarter the length of fourth ray; tips of pelvic-fin rays not ending in fleshy pads; caudal fin rounded; branched caudal-fin rays 15; segmented caudal-fin rays 17.
Scales: Scales on head and trunk absent.
Head: Jaw extending posteriorly to a vertical at anterior of eye; anterior nare an elongate narrow tube; posterior nare an opening with raised rim; no cephalic lateralis pores on head or preopercle; eyes 30.1% HL, dorsolateral, extending above head profile; interorbital narrow, 7.09% HL; snout profile steep; operculum opening extending slightly wider than vertical of pectoral-fin base; teeth in upper jaw in three or four rows, teeth in outer row enlarged, extending along most of premaxilla, teeth in inner rows small and more numerous; teeth in lower jaw in three or four rows, teeth in outermost and innermost rows slightly enlarged, middle rows smaller and more numerous, two enlarged recurved canines in inner row at midpoint of dentary.
Genitalia: Male with conical pointed papilla, no melanophores present; female unknown.
Colour in life ( Fig. 20 View Figure 20 ): Head and body translucent white (with a yellow cast to body after death), with distinct thin yellow – brown bars; head with four distinct bands radiating from eye; first band, with few melanophores, extending from anterior of eye forwards and down across both jaws; second band, with numerous melanophores, extending posteroventral corner of eye onto middle of preopercle; third band with many melanophores and dark edges, originates at posterodorsal corner of eye, extending posteriorly where it meets with corresponding band from opposite side of head, forming U – shape across the top of head; fourth band a short thin yellow interorbital bar connecting the anterodorsal corners of the two eyes; snout light yellow with a few scattered melanophores; trunk with five narrow, yellow – brown bars with scattered melanophores, densest along edges; first trunk bar across nape, connecting the upper corners of the pectoral fins; second trunk bar originating under dorsal-fin spines 4 – 6, extending to abdomen; third trunk bar extending from anterior of second dorsal fin to abdomen; fourth trunk bar originating under posterior one-third of second dorsal fin; fifth trunk bar over mid-caudal peduncle; first dorsal fin with bright-white stripe along outer edge and three yellow – brown bands; lower band on first dorsal fin dark yellow – brown, extending from lower anterior edge of fin to midpoint of base, where it joins the first body bar; second band on first dorsal fin extending diagonally from near mid-point of first spine to near posterior of fin base, yellow – brown with dense melanophores at front, broken into spots posterior to fifth spine; third and outermost band on first dorsal fin broad, yellow, with dark melanophores concentrated along first spine, colour becoming lighter posteriorly; second dorsal fin translucent white, with broad bright-white dorsal edge, with a row of yellow – brown spots under the rear third of that white band, and four yellow – brown transverse bands with numerous melanophores; first band extending only from first to third spine before coalescing with the third trunk bar, second band extending from first spine to seventh ray before coalescing with the fourth bar on trunk; third and fourth bands narrow, extend entire width of fin; caudal fin translucent white with rear margin broadly bright white, and with four thin yellow – brown vertical bars with numerous melanophores, bars equally spaced, anterior bar at base of caudalfin rays; lower one-fifth of caudal fin yellow with black margin; pectoral fin translucent to white on lower two-thirds of fin, bright yellow on upper onethird, with bright-yellow band coalescing with the first trunk bar; anal fin translucent with dark distal margin; anal fin translucent with blackish outer margin, rear edge yellowish; pelvic fin colour not known.
Colour in preservation ( Fig. 21 View Figure 21 ): Background colour of body and head yellowish pale; all bright-yellow markings from life still evident as dark concentrations of melanophores, including dark streak on upper third of pectoral fin, and dark stripes on head, nape, body, and dorsal and caudal fins; anal fin with dark posterior margin; pelvic fin immaculate.
Sensory papillae ( Fig. 22 View Figure 22 ): A transverse pattern with rows 1, 2, and 3/4 extending most of the distance from orbit to row d; row 5s/5i connected and at the anterior of row b; row b short, composed of four papillae; row d short, about four papillae located midway between rows 1 and 5s/5i; interorbital papillae row pc’ with two papillae present.
Vertebral skeleton: Dorsal pterygiophores formula 3 – 221110; one anal-fin pterygiophore inserted anterior to first haemal arch; second neural arch expanded, forked at tip; hypurals 1 and 2 fused to hypurals 3 and 4 along half of their length; 27 vertebrae – 11 precaudal and 16 caudal.
Habitat: The only known specimen was found inside a glass bottle collected along a sandy slope between 114 and 137 m depth. There is no additional information on the natural microhabitat of this species.
Distribution: Known only from the type location off Bonaire.
Etymology: Named after Laura Albini, wife of Adriaan ‘Dutch’ Schrier, the owner of Substation Curacao, through whose efforts new, tropical, deep- water species are being discovered. Laura has generously fed and hosted numerous researchers during their visits to Curacao.
Comparison: Psilotris laurae sp. nov. is the only deep-water species of Psilotris known at this time, and the only species with one anal-fin pterygiophore inserted anterior to the haemal spine. It can be distinguished from all other Psilotris species except Psilotris kaufmani by possessing a bicoloured pectoral fin. Psilotris laurae sp. nov. differs from Psilotris kaufmani in having distinct narrow vertical bars on the body and caudal fin versus small round spots and mottling ( Fig. S3D View Figure 3 ). The distinct vertical bars also serve to separate Psilotris laurae sp. nov. from the morphologically similar Varicus decorum sp. nov., which possess distinct yellow circles and spots along the side of the body and dorsal surface. In addition, Psilotris laurae has pelvic-fin rays 1 – 4 branched to the tips, whereas rays 1 – 4 are either unbranched or branched and refused as fleshy pads in Varicus decorum sp. nov.


| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Psilotris
| Tornabene, Luke, Van Tassell, James L., Gilmore, Richard G., Robertson, David Ross, Young, Forrest & Baldwin, Carole C. 2016 |
Carrigobius
| VAN TASSELL, TORNABENE & GILMORE 2016 |
Cryptopsilotris
| Tornabene & Van Tassell & Gilmore & Robertson & Young & Baldwin 2016 |
Carrigobius
| VAN TASSELL, TORNABENE & GILMORE 2016 |
Cryptopsilotris
| Tornabene & Van Tassell & Gilmore & Robertson & Young & Baldwin 2016 |
Psilotris amblyrhynchus
| Smith & Baldwin 1999 |
Psilotris amblyrhynchus
| Smith & Baldwin 1999 |
Psilotris amblyrhynchus
| Smith & Baldwin 1999 |
Psilotris amblyrhynchus
| Smith & Baldwin 1999 |
Psilotris bohlkei
| Greenfield 1993 |
Psilotris celsa
| Bohlke 1963 |
Psilotris batrachoides
| Bohlke 1963 |
Psilotris batrachodes
| Bohlke 1963 |
Psilotris batrachodes
| Bohlke 1963 |
Psilotris celsa
| Bohlke 1963 |
Psilotris batrachodes
| Bohlke 1963 |
Psilotris
| GINSBURG 1953 |
Psilotris
| GINSBURG 1953 |
Psilotris
| GINSBURG 1953 |
Psilotris
| GINSBURG 1953 |
Psilotris
| GINSBURG 1953 |
Psilotris
| GINSBURG 1953 |
Psilotris
| GINSBURG 1953 |
Chriolepis
| GILBERT 1892 |