
Galathea guttata Osawa, 2004
|
publication ID |
https://doi.org/10.12782/specdiv.25.295 |
|
persistent identifier |
https://treatment.plazi.org/id/03A0E24A-633C-FFD2-FEF7-FC51054C3F53 |
|
treatment provided by |
Felipe |
|
scientific name |
Galathea guttata Osawa, 2004 |
| status |
|
Galathea guttata Osawa, 2004 View in CoL ( Fig. 2H View Fig )
Galathea sp. B : Kato and Okuno 2001: 88, unnumbered fig.
Galathea guttata Osawa, 2004: 93 View in CoL , figs 1, 2, 3A, B ( type locality: Kume Island , Ryukyu Islands, Japan; 18 m); Baba et al. 2008: 69; Dong and Li 2010: 11 View Cited Treatment , fig. 6; Macpherson and Robainas-Barcia 2015: 133 View Cited Treatment .
Material examined. Tottori Prefecture. Off Tajiri, Iwami ( 35°59.1′N, 134°31.0′E), 12 m, SCUBA, 11 October 2017, coll GoogleMaps . Y GoogleMaps . Ota, 1 male (pcl 3.1 mm), TRPM-785.
Distribution. Japan, South China Sea, Papua New Guinea, and New Caledonia; depths of 1–54 m ( Macpherson and Robainas-Barcia 2015). Japanese records include the Ryukyu Islands ( Okinawa and Kume Islands), Izu Islands (Hachijo Island), and Tottori Prefecture ( Kato and Okuno 2001, as Galathea sp. B ; Osawa 2004; present study). The present specimen reveals the occurrence of this species in the Sea of Japan for the first time.
Habitat. Rock; dead and living coral; and coarse mud, sand and shell ( Osawa 2004; Dong and Li 2010). The present specimen was collected from the underside of a rock.
Remarks. This galatheid can be regarded as a tropical shallow-water species, estimated from the known distribution. The uninterrupted mesogastric ridge between the first branchial marginal spines on the carapace and two rounded white spots at the base of the cheliped fingers are diagnostic for G. guttata , and these characters immediately distinguish the species from other local congeners such as G. orientalis Stimpson, 1858 and G. peitho Macpherson and Robainas- Barcia, 2015, known from Tottrori and/or Shimane Prefectures ( cf. Osawa et al. 2014b, 2015, 2018).
Family Munididae Ahyong, Baba, Macpherson, and Poore, 2010 Munida agave Macpherson and Baba, 1993 ( Figs 2I View Fig , 3D View Fig )
Munida agave Macpherson and Baba, 1993: 387 , figs 1, 2 ( type locality: Philippines, MUSORTOM 2 stn 33, 13°32.3′N, 121°07.5′E; 130– 137 m.); Macpherson 1997: 603; Baba 2005: 89, 258; Baba et al. 2008: 84 View Cited Treatment (compilation).
Material examined. Tottori Prefecture. Off Nagaobana ( 35°55′N, 134°00′E), Tottori: 100 m, beam trawl, 15 May 2018, coll GoogleMaps . Y GoogleMaps . Ota , 2 males (pcl 5.9, 6.0 mm), TRPM-786; 70 m, beam trawl, 13 July 2018, coll . Y . Ota , 1 ovigerous female (pcl 7.5 mm), TRPM-787; 100 m, beam trawl, 13 July 2018, coll . Y . Ota , 1 ovigerous female (pcl 7.4 mm), TRPM- 788; 70 m, beam trawl, 17 April 2019, coll . Y . Ota, 1 female (pcl 5.3 mm), TRPM-789.
Distribution. Japan, the Philippines, and Indonesia; depths of 89–549 m ( Baba et al. 2008). Japanese records include Sagami Bay, and Tottori and Kagoshima Prefectures ( Macpherson and Baba 1993; Baba 2005; present study). The present specimens reveal the first record of this species from the Sea of Japan.
Habitat. Hard bottom, rock ( Baba 2005). The present specimens were collected from the muddy sand substrate.
Remarks. The present specimens of Munida agave were found together with another congener, M. pherusa Macpherson and Baba, 1993 , in the same trawl catch collected from off Nagaobana, at depths of 80– 100 m. The latter species has been already recorded from the Sea of Japan, off Yamaguchi Prefecture ( Komai et al. 2002). The two species are separated for each other by characters discussed by Macpherson and Baba (1993), as well as the fresh coloration. The bases of the rostrum and cheliped fingers have a white patch respectively in M. pherusa unlike in M. agave ( Fig. 3D–F View Fig ).
Paramunida tricarinata ( Alcock, 1894) [New standard Japanese name: Mitsutoge-zarazara-chu-koshiori-ebi] ( Figs 3G View Fig , 4 View Fig )
Restricted synonymy.
Munida tricarinata Alcock, 1894: 324 ( type locality: Andaman Sea, 205 m); Alcock and Anderson 1895: pl. 12, fig. 1.
Paramunida scabra ( non Henderson, 1885): Wu et al. 1998: 145, figs 41, 42G; Macpherson 1993: 462 (in part).
Paramunida tricarinata: Baba 2005: 304 ; Baba et al. 2008: 175 (list of occurrences); Baba et al. 2009: 283, figs 260, 261; Cabezas et al. 2010: 56 View Cited Treatment , figs 15H, 18I; McCallum et al. 2016: 216 View Cited Treatment , figs 2B, C, 8.
Material examined. Tottori Prefecture. Off Nagaobana ( 35°55′N, 134°00′E), Tottori: 100 m, beam trawl, 15 May 2018, coll GoogleMaps . Y GoogleMaps . Ota, 1 male (pcl 6.6 mm), 1 ovigerous female
(pcl 7.9 mm), TRPM-790; 100 m, beam trawl, 20 August 2018, coll. Y. Ota, 1 female (pcl 6.2 mm), 1 ovigerous female (pcl. 5.5 mm), TRPM-791; 120 m, beam trawl, 20 August 2018, coll. Y. Ota, 1 female (pcl 6.3 mm), TRPM-792.
Distribution. Taiwan, Philippines, Andaman Sea, northwestern Australia, Maldives Islands, and Arabian Sea; depths of 187–384 m ( McCallum et al. 2016). Newly record- ed from Japanese waters.
Habitat. No previous records on the collection substrate, but vast majority of the previous specimens were collected by trawl, suggesting that they were probably on soft bottoms. The present specimens were collected from muddy sand substrates.
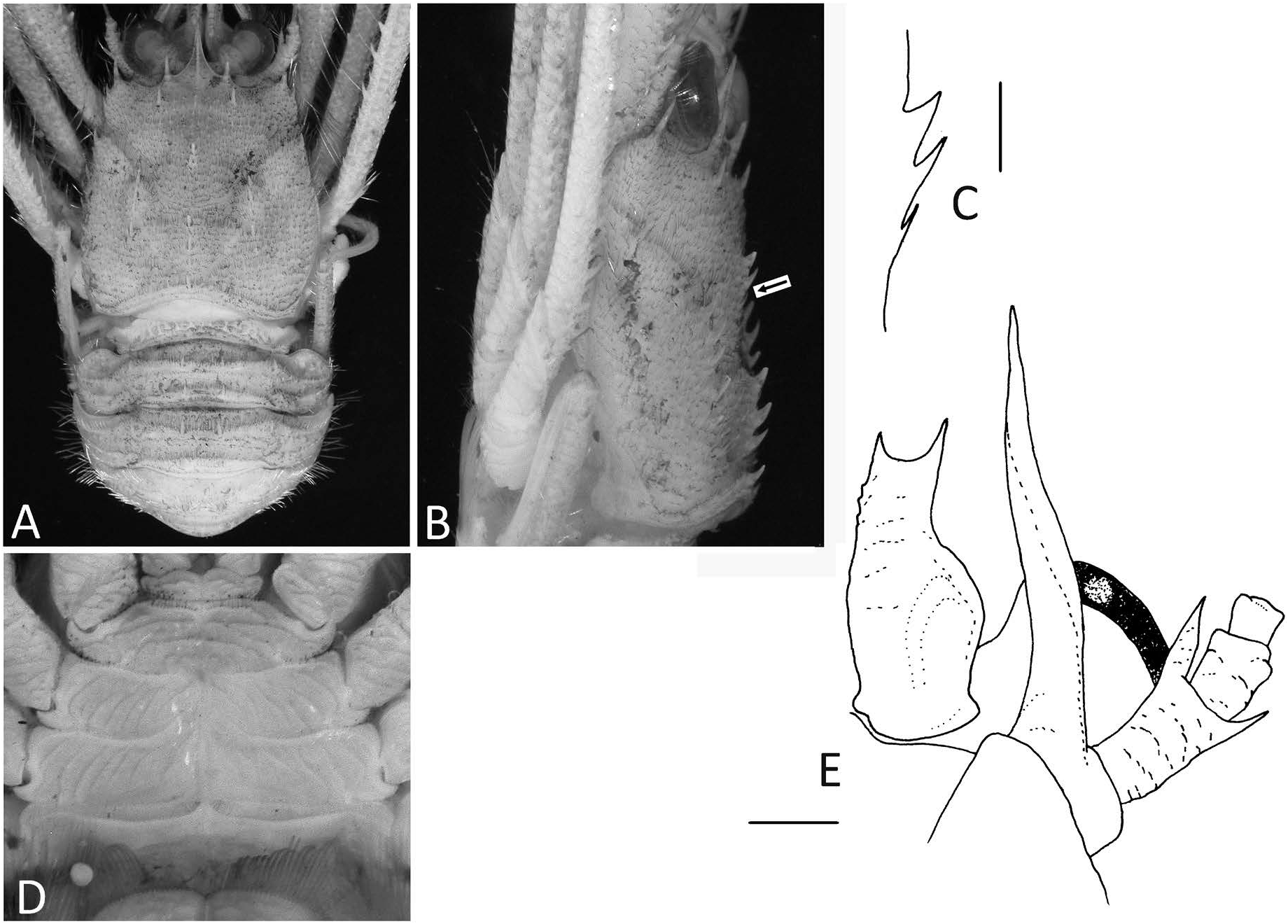
Remarks. The present material from the Sea of Japan agrees well with Taiwan and Philippine specimens of P. tricarinata reported by Baba et al. (2009) and Cabezas et al. (2010) in the following diagnostic features ( Fig. 3 View Fig ): the rostral spine is larger than the supraocular spines; the carapace has a row of three spines in the midline of the mesogastric region and tufts of long and dense setae along the anterior branch of the cervical groove; the sternal plastron has numerous striae; the anterior prolongation of the antennal article 1 is spiniform and does not have a row of spines on the dorsal margin; the antennal article 2 is slightly longer than broad, the distomesial spine is spiniform and slightly overreaches the end of the antennal peduncle at most (in the present material, it reaches the end of the antennal peduncle), the distolateral spine never reaches the end of that article; and the antennal article 3 is slightly longer than broad. Unlike these specimens from Japan, Taiwan, and the Philippines, the antennal article 2 of the north-western Australian material reported by McCallum et al. (2016) has the distomesial spine extending far beyond the end of the antennal peduncle and the distolateral spine reaching or slightly exceeding the end of the article 3, although the Tamura-Nei genetic divergence using the mitochondrial ND1 and 16S rRNA genes between specimens of P. tricarinata from north-western Australia and Taiwan is lower than those between sister taxa (0.008±0.004 vs. 0.018±0.004 between P. ascella McCallum, Cabezas and Andreakis, 2016 and P. tricarinata ; McCallum et al. 2016).
The size of the median spines in the mesogastric region of the carapace may vary in P. tricarinata . In the present material, the three spines on the mesogastric median row decrease in size posteriorly; the anterior two spines are strong and the third spine is smaller than the second or greatly reduced in size ( Fig. 4B, C View Fig ), as illustrated for a Philippine specimen by Cabezas et al. (2010: fig. 15H). In the illustrated specimens from Taiwan and north-western Australia ( Baba et al. 2009: fig. 261a; McCallum et al. 2016: fig. 8A, B), the spines are subequal in size and the third spine is rather large. The anterior prolongation of the antennal article 1 is also proportionally shorter in the present material than the previously reported specimens. In the material examined, it overreaches the distal margin of the antennular article 1 by one third ( Fig. 4E View Fig ), instead of about half of its length in other described/illustrated specimens ( Cabezas et al. 2010: fig. 18I; McCallum et al. 2016: fig. 8D).
Although literature records of P. tricarinata include western Indian Ocean localities ( Laurie 1926; Tirmizi 1966, as Munida Leach, 1820 ; Baba 1990; Macpherson 1993), identities of these specimens were questioned by Cabezas et al. (2010).
In Japanese waters, three congeneric species, P. leptotes Macpherson and Baba, 2009 ( Baba 1982, 1986; as Munida proxima Henderson, 1885 ), P. scabra (Henderson, 1885) , P. stichas Macpherson, 1993 , are known ( cf. Baba 2005; Macpherson and Baba 2009). Paramunida tricarinata is the fourth species of the genus recorded from Japan.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Galathea guttata Osawa, 2004
| Osawa, Masayuki & Ota, Yuzo 2020 |
Paramunida tricarinata :
| McCallum, A. W. & Cabezas, P. & Andreakis, N. 2016: 216 |
| Cabezas, P. & Macpherson, E. & Machordom, A. 2010: 56 |
| Baba, K. & Macpherson, E. & Lin, C. - W. & Chan, T. Y. 2009: 283 |
| Baba, K. & Macpherson, E. & Poore, G. C. B. & Ahyong, S. T. & Bermudez, A. & Cabezas, P. & Lin, C. - W. & Nizinski, M. & Rodrigues, C. & Schnabel, K. E. 2008: 175 |
| Baba, K. 2005: 304 |
Galathea guttata
| Macpherson, E. & Robainas-Barcia, A. 2015: 133 |
| Dong, C. & Li, X. 2010: 11 |
| Baba, K. & Macpherson, E. & Poore, G. C. B. & Ahyong, S. T. & Bermudez, A. & Cabezas, P. & Lin, C. - W. & Nizinski, M. & Rodrigues, C. & Schnabel, K. E. 2008: 69 |
| Osawa, M. 2004: 93 |
Galathea sp. B
| Kato, S. & Okuno, J. 2001: 88 |
Munida agave
| Baba, K. & Macpherson, E. & Poore, G. C. B. & Ahyong, S. T. & Bermudez, A. & Cabezas, P. & Lin, C. - W. & Nizinski, M. & Rodrigues, C. & Schnabel, K. E. 2008: 84 |
| Baba, K. 2005: 89 |
| Macpherson, E. 1997: 603 |
| Macpherson, E. & Baba, K. 1993: 387 |
Munida tricarinata
| Alcock, A. 1894: 324 |