
Cnemidocarpa bythia ( Herdman, 1881 )
|
publication ID |
https://doi.org/ 10.1080/00222930400026969 |
|
persistent identifier |
https://treatment.plazi.org/id/03A10234-FFAC-2F69-9AED-29B1FD90FC7B |
|
treatment provided by |
Felipe |
|
scientific name |
Cnemidocarpa bythia ( Herdman, 1881 ) |
| status |
|
Cnemidocarpa bythia ( Herdman, 1881) View in CoL
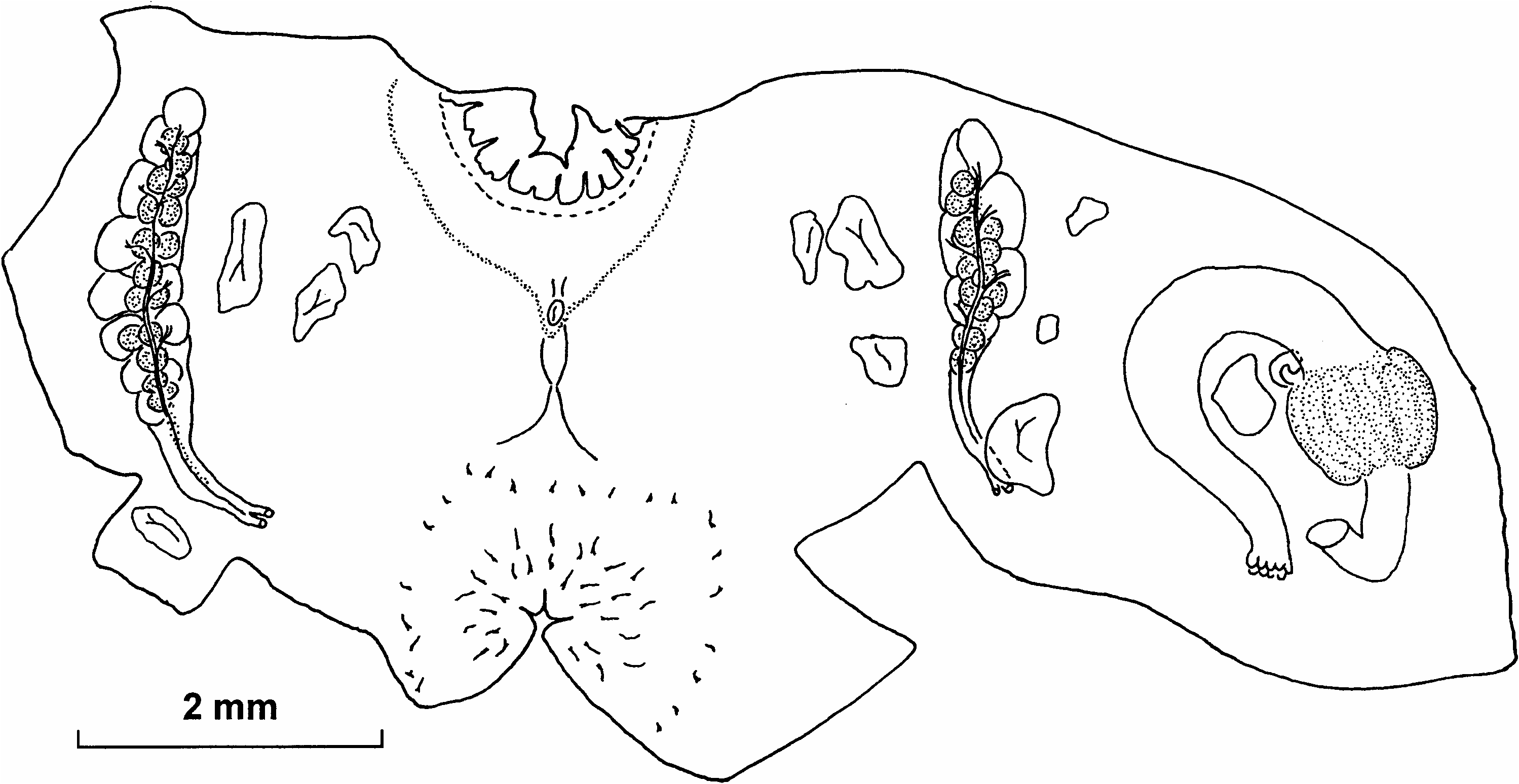
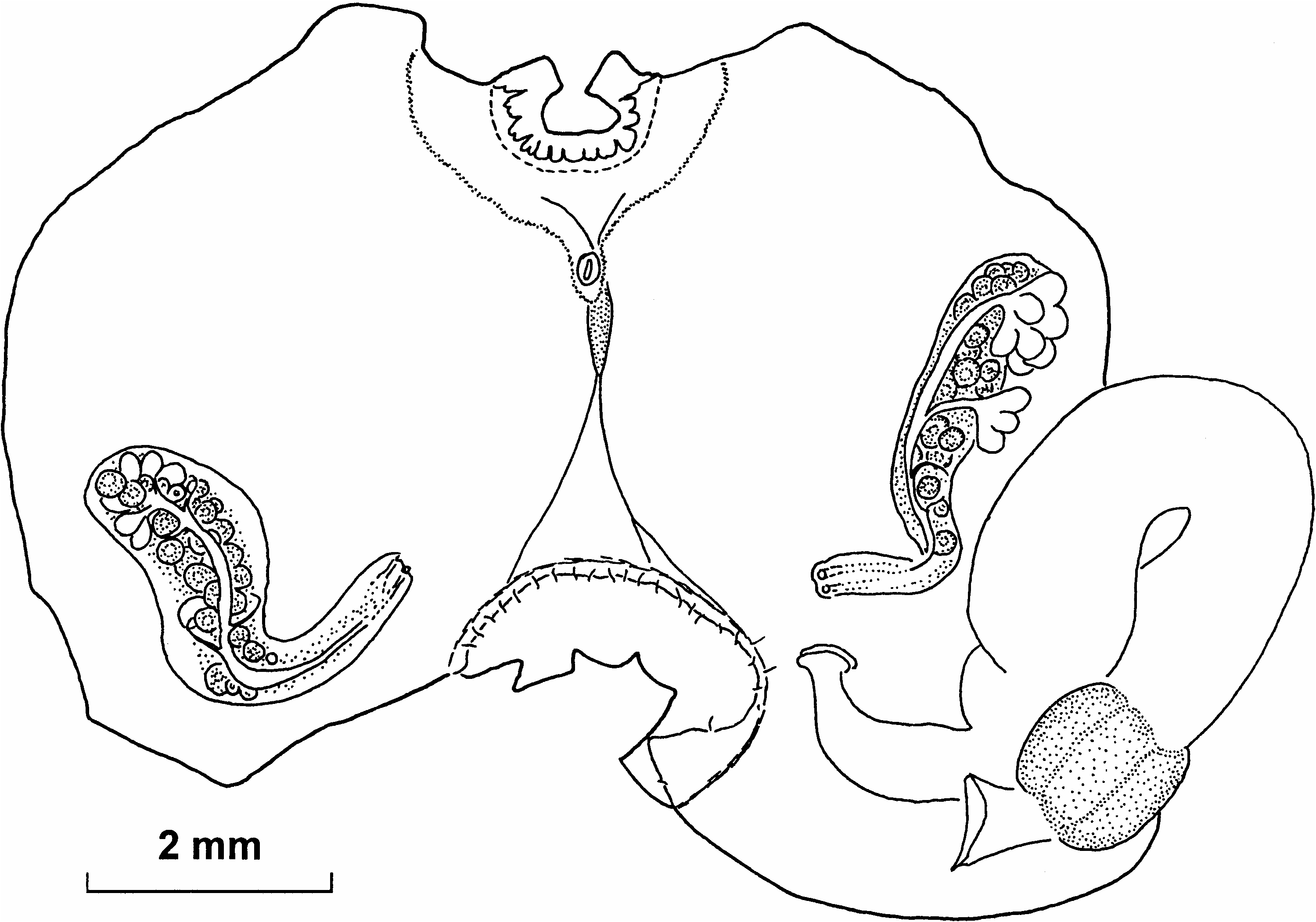
( Figures 2 View Figure 2 , 3 View Figure 3 )
Styela bythia Herdman 1881, p 63 View in CoL ; 1882, p 151. Cnemidocarpa bythia: Millar 1959, p 194 View in CoL ; Monniot and Monniot 1973, p 432; 1974, p 752; 1977, p 313; 1982, p 113; 1985b, p 27; Monniot 1998, p 552; Sanamyan and Sanamyan 2002, p 340.
Material examined
St. 4226, 3600–3630 m, five specimens; st. 4254, 3796– 3780 m, one specimen.
Description
Unlike many small deep-water Styelidae , that are almost impossible to distinguish without dissection, this species has constant and distinctive external features. The body is hemispherical or short and cylindrical, 3–4 mm in diameter and height and always is attached to solid objects such as stones or polychaete tubes. Some specimens have test hairs, on the basal part of the body, only in the places in contact with the sand or mud, where the specimen is attached to an object smaller than the base of the body. Otherwise the test has neither the hairs nor attached sand, although it is covered by crowded minute (0.025 mm) papillae or granules. Sessile apertures are on opposite sides of the upper surface.
Internal features are also stable. The number of branchial tentacles in the present specimens (13 or 14) is almost exactly the same as in South Atlantic specimens ( Sanamyan and Sanamyan 2002). The atrial velum is short and possibly reduced in places, but the ring of atrial tentacles is complete, although they are relatively sparse. The test lining the internal surface of the atrial siphon has short papillae different in structure from the atrial tentacles and from the papillae on the remainder of the test; they are not present on the test lining the branchial siphon. The prepharyngeal band makes a shallow V around the minute dorsal tubercle, which has a simple vertical slit. The neural ganglion is halfway between the siphons. The dorsal lamina has numerous languets. Only three folds of the branchial sac are well formed, the fourth (most ventral) fold is almost flat. The branchial formula of the figured specimen is: E10(10)(10)(15)DL(12)(10)(11)7E.
About 13 transverse rows of short stigmata, some rows are crossed by parastigmatic vessels. On the left the most ventral row contains oblique stigmata, but protostigmata were not detected. The stomach is distinctly demarcated from the oesophagus and the gut, and has a large curved caecum; the number of longitudinal folds, about 14, is higher than reported previously for South Atlantic specimens ( Sanamyan and Sanamyan 2002). The anal border is distinctly lobed. One elongate gonad with long gonoducts is on each side of the body. The testes are more mature than the ovaries: the right gonad has more than 10 large male follicles in two conspicuous rows between the ovary and the body wall and projecting from the sides of the ovary, the left gonad has six similarly placed follicles. Several, mostly large, endocarps are on each side of the body around the gonads, and one is in the gut loop.
Remarks
The holotype of this species is from the waters south of Australia, it is 2 cm long (much larger than the Atlantic specimens), and has ‘‘extremely numerous’’ longitudinal branchial vessels ( Herdman 1882, p 151). Such large specimens were also recorded from the Pacific sector of Antarctica, but never in the Atlantic ( Monniot and Monniot 1982). The specimens from the Tasman Sea and Kermadec Trench ( Millar 1959) have a plain-edged anal margin and a little more numerous longitudinal branchial vessels. As suggested previously ( Sanamyan and Sanamyan 2002), south-west Pacific specimens with a smooth anal border, and specimens from the Atlantic with a lobed anal margin may belong to different species. This is supported by the present specimens. Sanamyan and Sanamyan (2002) suggested that the presence of protostigmata separate Atlantic and Tasman Sea species. However, protostigmata occur only occasionally in this species and are not a valid distinguishing character and at this stage the records appear to represent a single species. Nevertheless, most recent published records lack descriptions and further data may indicate that the Atlantic specimens are a separate species.
The species has been recorded in the North and South Atlantic, south from Kerguelen Island, south from Australia, the Ross Sea, Kermadec Trench and northern Peru.
Polycarpa albatrossi ( Van Name, 1912) View in CoL
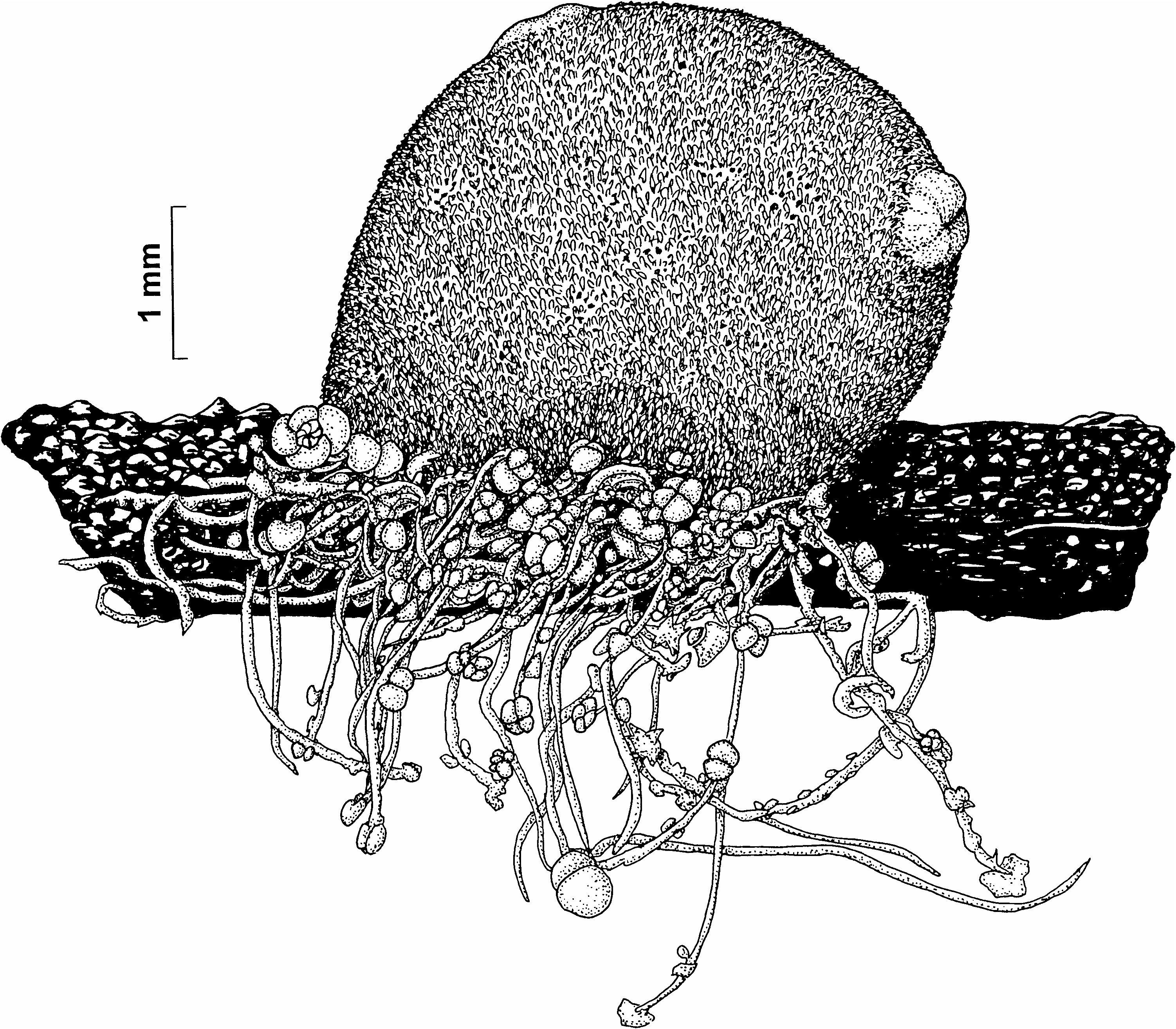
( Figures 4 View Figure 4 , 5 View Figure 5 )
Pandocia albatrossi Van Name 1912, p 579 View in CoL .
Polycarpa albatrossi: Van Name 1945, p 256 View in CoL ; Monniot and Monniot 1968, p 10; 1970, p 320; 1974, p 750.
Not Polycarpa albatrossi: Millar 1959, p 195 View in CoL . Material examined
St. 4254, 3796– 3780 m, 12 specimens.
Description
The specimens are almost globular or slightly flattened dorso-ventrally, 8–10 mm in diameter, not attached. Thin test hairs are not strongly branched and in general do not form such a dense mat as on some other deep-water species. Sand grains and foraminiferans firmly attached to the test hairs form a solid layer over the sides of the body, but are sparse on the proximal (ventral) side and almost absent on the dorsal side around and between the siphons. The low wide siphons on the upper surface are entirely free from foreign matter. Siphons are close together but not in contact with each other, they are always separated by a narrow area covered by sand. There are about 45 large simple branchial tentacles, sometimes projecting from the branchial siphon. The prepharyngeal band is a high and thick lamella with a shallow groove on the free edge, making almost a perfect circle without a dorsal V. The small dorsal tubercle has a transverse or C-shaped slit. The neural ganglion is just posterior, and close, to the dorsal tubercle, halfway between the siphons. Slender atrial tentacles arise from the base of the atrial velum, but not from its margin. The dorsal lamina is plain edged. Each side of the branchial sac has four unequal folds: the first (dorsal) and the third folds are always high, while the second and the fourth (ventral) folds are shallow and sometimes flat, indicated only by a group of longitudinal vessels. Only occasionally is a longitudinal vessel between the folds. Branchial formula of three specimens (high folds are in bold): E1(4)1(9)(7)(11)DL(10)(6)(12)(5)E; E(6)(11)(9)(12)DL(10)(9)(11)(6)E; E(5)(9)(7)(10)DL(9)1(6)(9)(5)E
Stigmata are in about 16–19 transverse rows, many crossed by parastigmatic vessels. All are longitudinal (protostigmata are not present). The stomach has about 14 well-marked longitudinal folds and a large curved caecum. The anal margin is lobed. Usually three gonads are on each side, but one specimen had five gonads on the left side. Gonads on each side of the body are irregularly spaced, in a row halfway to three-quarters of the way down each side of the body, each gonad placed at right angles to the antero-posterior axis of the body, its short duct directed toward the atrial aperture ( Figure 4 View Figure 4 ). Mature gonads are 1.25– 1.5 mm in length, male and female openings are inconspicuous and almost sessile, not visible without staining. The female opening is terminal, the male opening is a short distance from it on the upper surface of the anterior half of the gonad. Male follicles numerous, forming clusters between the oocytes and the body wall, sometimes slightly projecting from the sides. Numerous endocarps distributed evenly on the whole inner body wall including the space inside the gut loop. The size of the endocarps varies from specimen to specimen; some individuals have endocarps larger than are shown in Figure 4A View Figure 4 .
Remarks
Only two descriptions of Polycarpa albatrossi exist: the original description based on several specimens recorded in the north-west Atlantic near the coast of the USA ( Van Name 1912, 1945), and a more recent redescription based on specimens collected almost exactly in the same location ( Monniot and Monniot 1968). The main features documented for the species are several, but not numerous, gonads (instead of giving the exact number, Van Name 1945 reported that gonads are not so numerous as in Polycarpa fibrosa , and Monniot and Monniot 1968 recorded only that the gonads are always more numerous on the right side than the left); endocarps are numerous, and only the first and third branchial folds are perfectly developed. The present specimens show some differences from the Monniot and Monniot (1968) description. They are generally more like the specimens described by Van Name, the number of longitudinal branchial vessels in our specimens and the specimens described by Van Name (1912) are twice that in the smaller specimens described by Monniot and Monniot (1968). The main difference is in the structure of the gonads which in Monniots’ specimens have only three large male follicles in each, while the present specimens always have numerous small male follicles. There are several other records of this species, but they lack descriptions, stating only that the specimens conform exactly to existing accounts and thus cannot be used to clarify the range of variation.
Unlike many other deep-water species Polycarpa albatrossi has a limited distribution and has been recorded only from the north-east Atlantic. Recorded depth range is 2496–3761 m.
Styela calva Monniot, Monniot and Millar, 1976 View in CoL
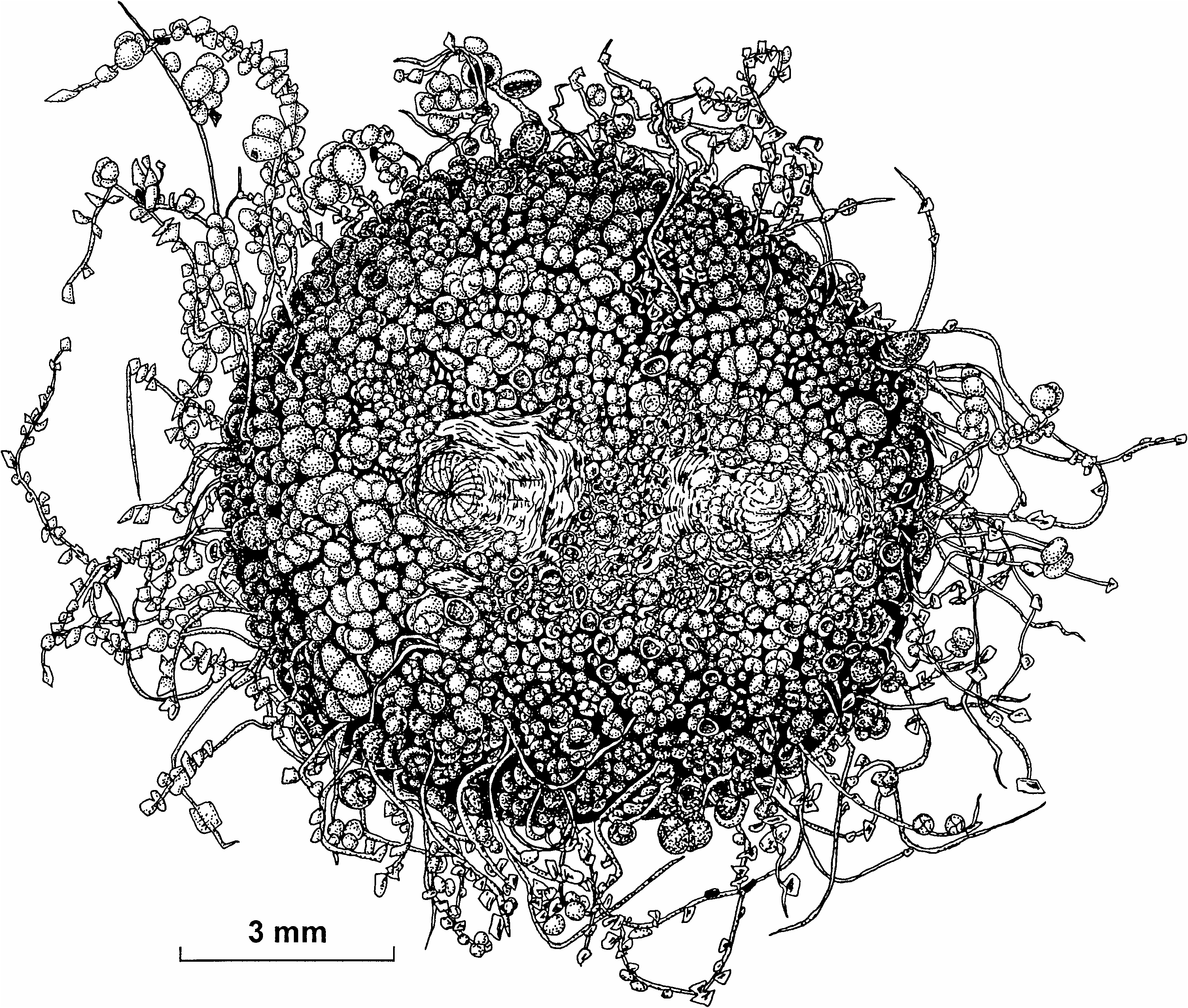
( Figures 6 View Figure 6 , 7 View Figure 7 )
Styela calva Monniot, Monniot and Millar, 1976, p 1196 View in CoL ; Millar 1982, p 172; Monniot and Monniot 1985a, p 296; 1985b, p 28; 1988, p 423.
Material examined
St. 4569, 3182– 3152 m, three specimens.
Description
The specimens are slightly flattened dorso-ventrally or laterally. Their greatest dimension is 4.5–6 mm. Test hairs with attached sand grains form a dense mat on the ventral half of the body, while the upper (dorsal) side is devoid of any outgrowths, although the whole body is covered by a solid layer of sand grains and foraminiferans. Apertures are widely separated, at opposite ends of the upper surface on inconspicuous sessile siphons. About 19 short branchial tentacles arise from the margin of a short branchial velum. The prepharyngeal band has a short, narrow but distinct dorsal V. The dorsal tubercle has a simple, vertical slit. The neural ganglion is just posterior and close to the dorsal tubercle, halfway between the siphons. The dorsal lamina is plain edged. The atrial velum, very short dorsally, has a large triangular laminar expansion on the left side, opposite to the intestine. The corresponding place on the right side of the atrial velum is damaged, but according to Monniot et al. (1976, Figure 15) this species has two triangular expansions on the ventral side, on the left and the right. Slender, short atrial tentacles are evenly distributed along the atrial velum. They were not found to be in two bunches opposite the genital opening as described by Monniot et al. (1976). Tentacles are not present on the triangular expansion of the velum. The branchial sac is damaged; apparently it has only three folds, the most ventral (fourth) fold being reduced. Each fold has about eight longitudinal vessels. The gut forms a closed narrow loop. The intestine is filled by fine particles and is very thick; the small, almost globular stomach has six to eight poorly defined longitudinal folds; and the anal border is bilobed and smooth. A pyloric caecum was not detected. One elongate gonad is on each side. Male follicles are in bunches between the ovary and the body wall and projecting slightly from the sides of the ovary. Gonoductal openings are close together, each at the end of short gonoducts at the distal end of each gonad. The gonoducts and the distal part of each gonad turn dorsally toward the atrial aperture, almost at right angles to its proximal end. Distinct endocarps are not present, but the pallial wall has a low, cushionlike delamination of the epithelium. Similar structures are probably present in the specimens described by Monniot et al. (1976, p 1197), since they wrote that ‘‘the body wall has a tendency to delaminate possibly as a result of fixation’’.
Remarks
The most significant distinctive features of this species are the absence of the endocarps and the characteristic shape of the atrial velum. The smooth bilobed anus and the ‘‘delaminated’’ cushions of the pallial epithelium are also characteristic. The latter feature is not a result of fixation as was supposed by Monniot et al. (1976). The original description of this species is based on many (about 40) specimens from the north-east and south-east Atlantic, and on one specimen from the Tasman Sea. The latter specimen, geographically distant from all others, was chosen as the holotype, although the reason for this selection is not explained. It is not known which specimen is figured in the original description, the Tasman Sea specimen or one from the Atlantic ( Monniot et al. 1976, Figure 15). Further records include specimens from the north-east, central and south-east Atlantic, north and central Indian Ocean.

| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Cnemidocarpa bythia ( Herdman, 1881 )
| Sanamyan, K. E. & Sanamyan, N. P. 2005 |
Styela calva
| Monniot C & Monniot F 1985: 296 |
| Millar RH 1982: 172 |
| Monniot C & Monniot F & Millar RH 1976: 1196 |
Polycarpa albatrossi:
| Millar RH 1959: 195 |
Polycarpa albatrossi:
| Monniot C & Monniot F 1968: 10 |
| Van Name WG 1945: 256 |
Pandocia albatrossi
| Van Name WG 1912: 579 |
Styela bythia
| Sanamyan K & Sanamyan N 2002: 340 |
| Monniot C 1998: 552 |
| Monniot C & Monniot F 1973: 432 |
| Millar RH 1959: 194 |
| Herdman WA 1881: 63 |