
Alteutha alsagopu, Gómez & Varela, 2013
|
publication ID |
https://doi.org/10.1080/00222933.2012.747634 |
|
persistent identifier |
https://treatment.plazi.org/id/03A1295F-FFE9-7A45-FE20-FCE0FC4B3EE9 |
|
treatment provided by |
Felipe |
|
scientific name |
Alteutha alsagopu |
| status |
sp. nov. |
Alteutha alsagopu sp. nov.
( Figures 1–9 View Figure 1 View Figure 2 View Figure 3 View Figure 4 View Figure 5 View Figure 6 View Figure 7 View Figure 8 View Figure 9 )
Material examined
One dissected female holotype (EMUCOP-050997-01), one male allotype preserved in ethanol (EMUCOP-010805-01), one adult, one CII and one CIII paratype (EMUCOP-010805-02) and one male paratype (EMUCOP-170801-41) preserved in ethanol, and two dissected male (EMUCOP-010805-03, EMUCOP-260298-02) and one dissected female (EMUCOP-260298-01) paratypes. Collected from Urías System ( Sinaloa State, NW Mexico) ( 23 ◦ 11 ′ 06 ′′ N, 106 ◦ 25 ′ 06 ′′ W); 5 September 1997, 26 February 1998, 17 August 2001, 1 August 2005; coll. S. Gómez ( 5 September 1997), E. Barranco Ramírez ( 26 February 1998), and S. Gómez and F. N. Morales-Serna ( 17 August 2001, 1 August 2005).
Type locality
Urías System ( Sinaloa State, NW Mexico) ( 23 ◦ 11 ′ 06 ′′ N, 106 ◦ 25 ′ 06 ′′ W) GoogleMaps .
Etymology
The species was named after the son of the senior author, Alfonso Samuel Gómez Puello, using the first two letters of his names (AlSaGoPu).
Description
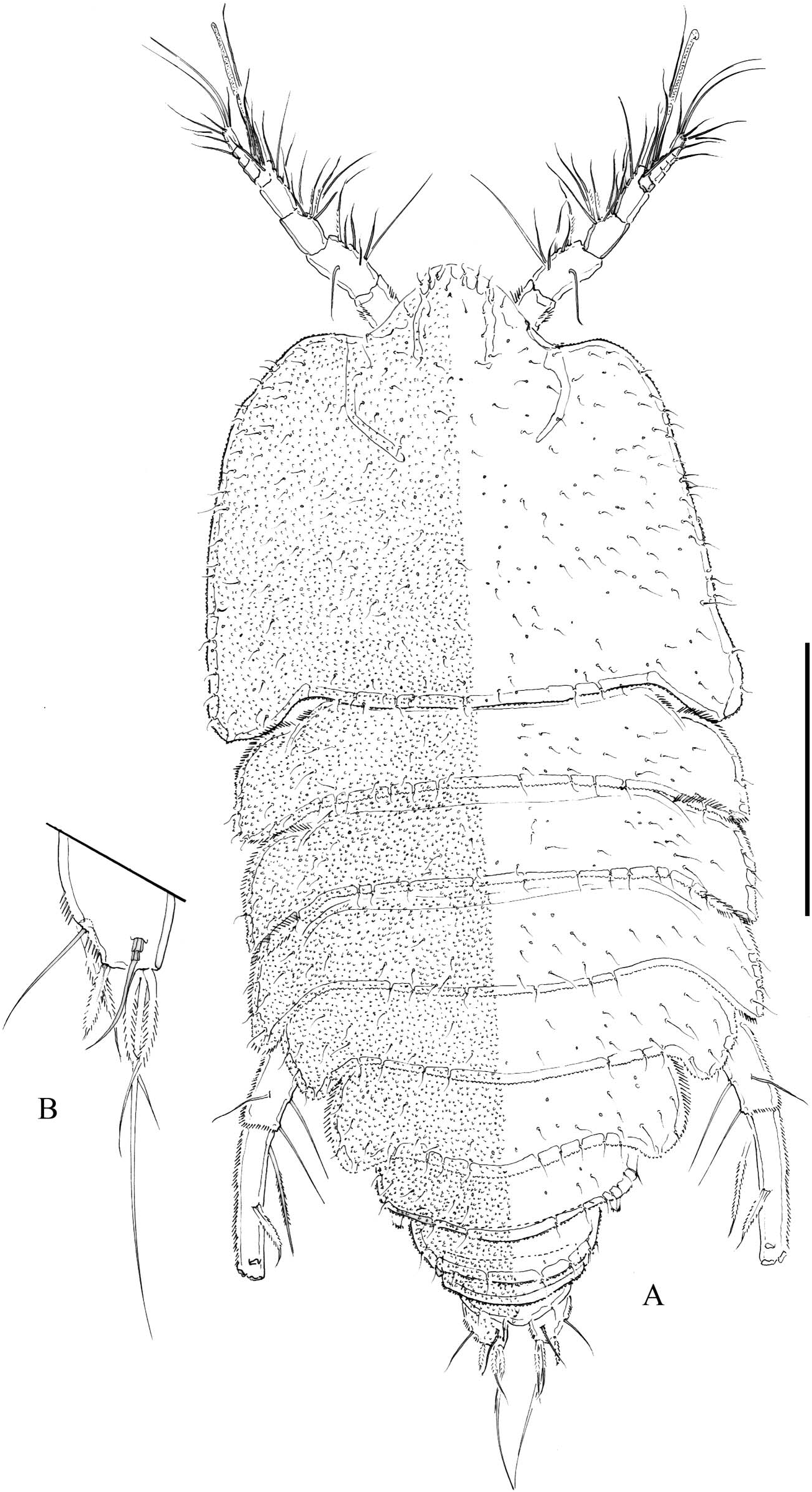
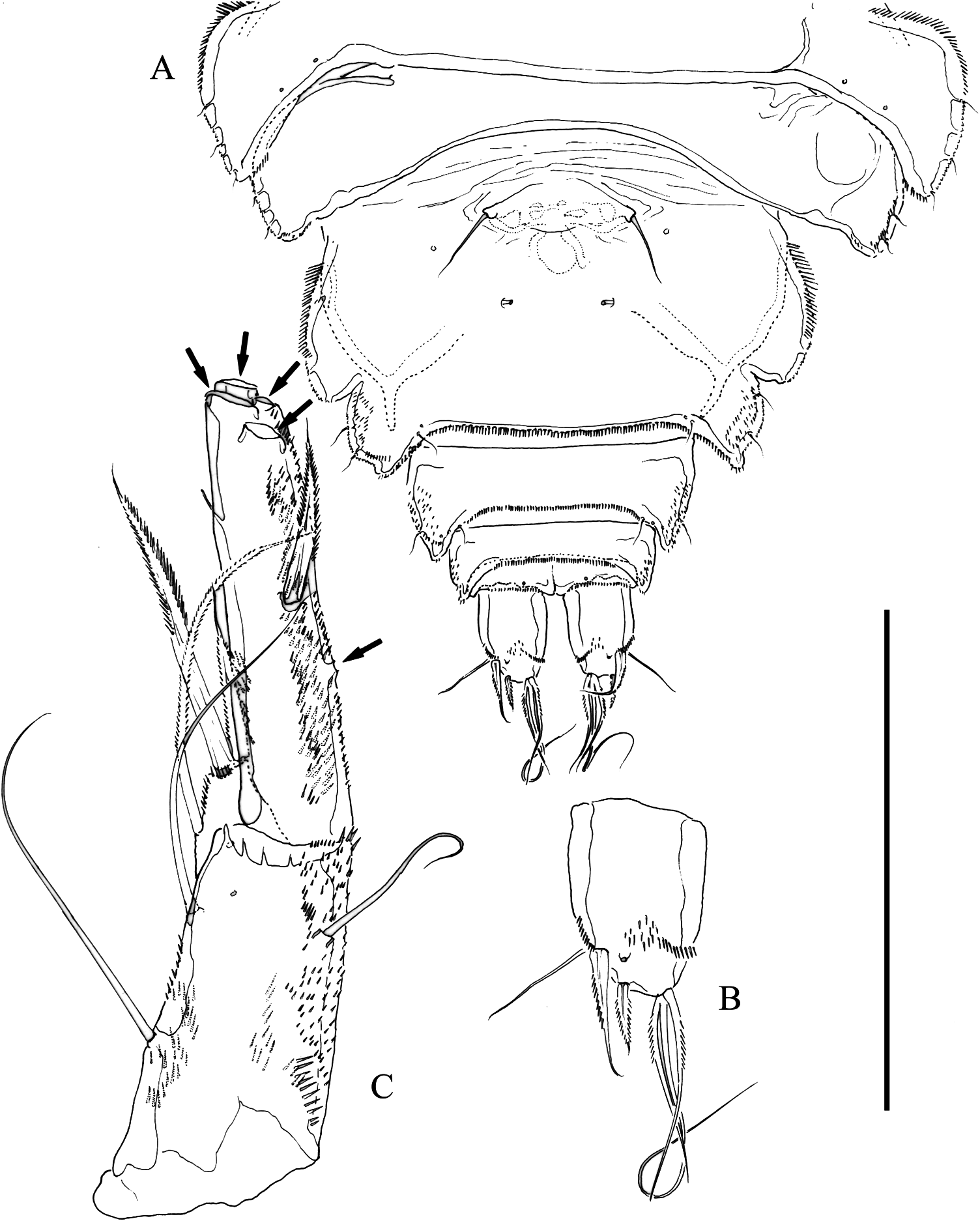
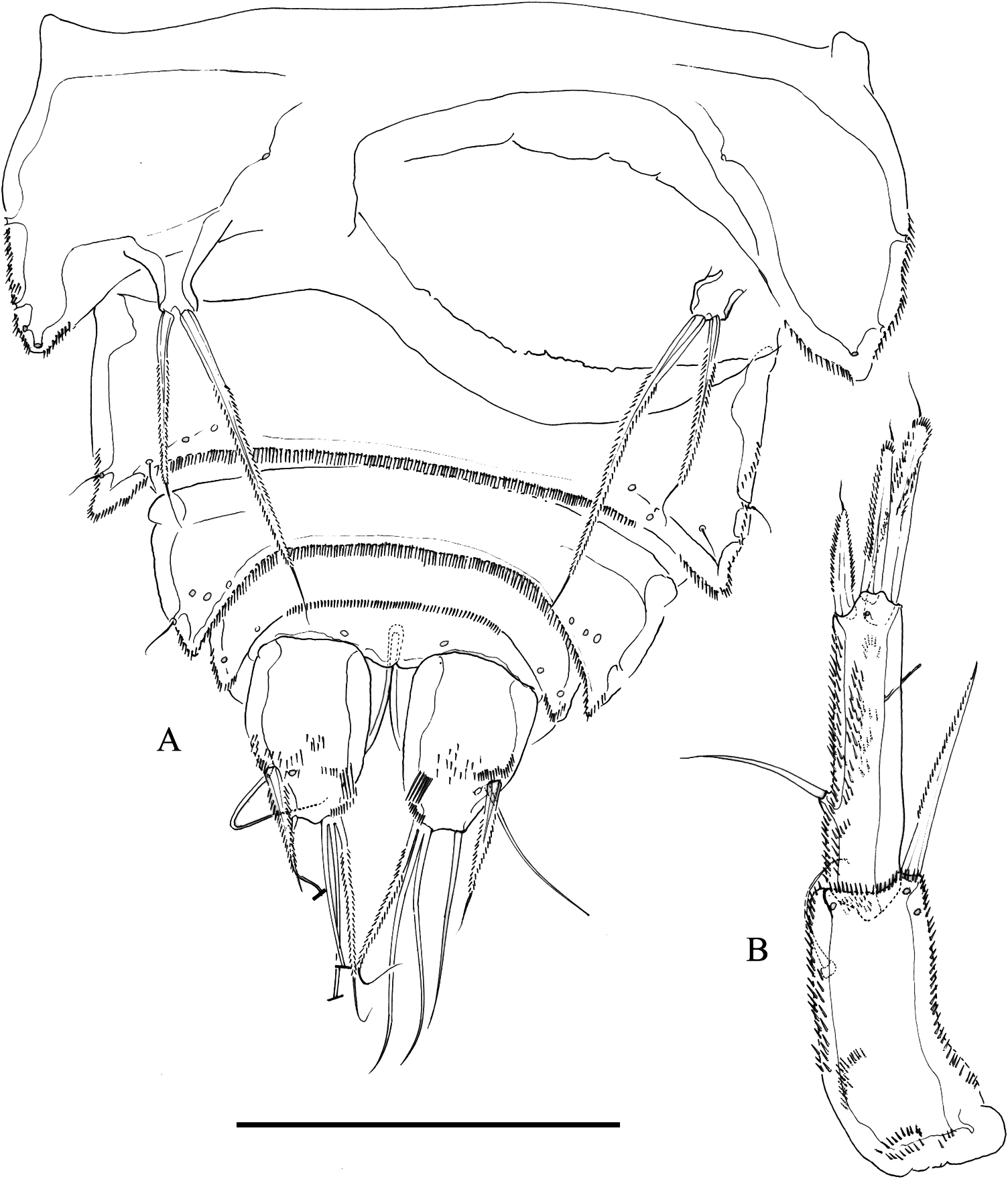
Female. Body ( Figure 1A View Figure 1 ) broad, flattened, somewhat arched along midline, tapering posteriorly, without pattern of chitinous thickenings. Total body length of holotype, 790 µm measured from tip of rostrum to posterior margin of caudal rami. Dorsal surface of cephalothorax, pro- and urosomites covered with minute denticles, with posterior margin minutely serrate; with sensilla as figured. Rostrum fused to cephalothorax, triangular, with rounded tip. Three free prosomites, P5-bearing somite, genital double-somite, and fourth urosomite with, fifth urosomite without sensilla. Genital double-somite partially fused dorsally, completely fused ventrally ( Figure 2A View Figure 2 ); second half of genital double-somite with spinular row along posterior margin; genital field situated proximally on first half of genital double-somite; each P6 represented by slen- der seta. Fourth, fifth and sixth urosomites as preceding somite ventrally. Caudal rami ( Figures 1B View Figure 1 , 2B View Figure 2 ) slightly tapering posteriorly, with spinules as figured; about 1.4 times as long as wide; with seven elements; seta I bipinnate, spine-like, nearly as long as ramus width; seta II a slender seta, as long as and dorsal to seta I; seta III bipinnate, spine-like, noticeably shorter than seta I and situated on distal outer corner; seta IV and V fused basally; seta IV bipinnate and about half the length of seta V; seta VI bipinnate, about as long as seta IV, situated on distal inner corner; seta VII situated rather distally, close to inner corner, somewhat shorter than seta II, biarticulated at its base.
Antennule ( Figure 3B View Figure 3 ) nine-segmented; first and second segments with spinules as figured; with several pinnate setae (most setae naked and slender). Armature formula as follows: I – (1); II – (12); III – (11 + ae); IV – (4 + ae); V – (2); VI – (4); VII – (2); VIII – (2); IX – (7 + ae).
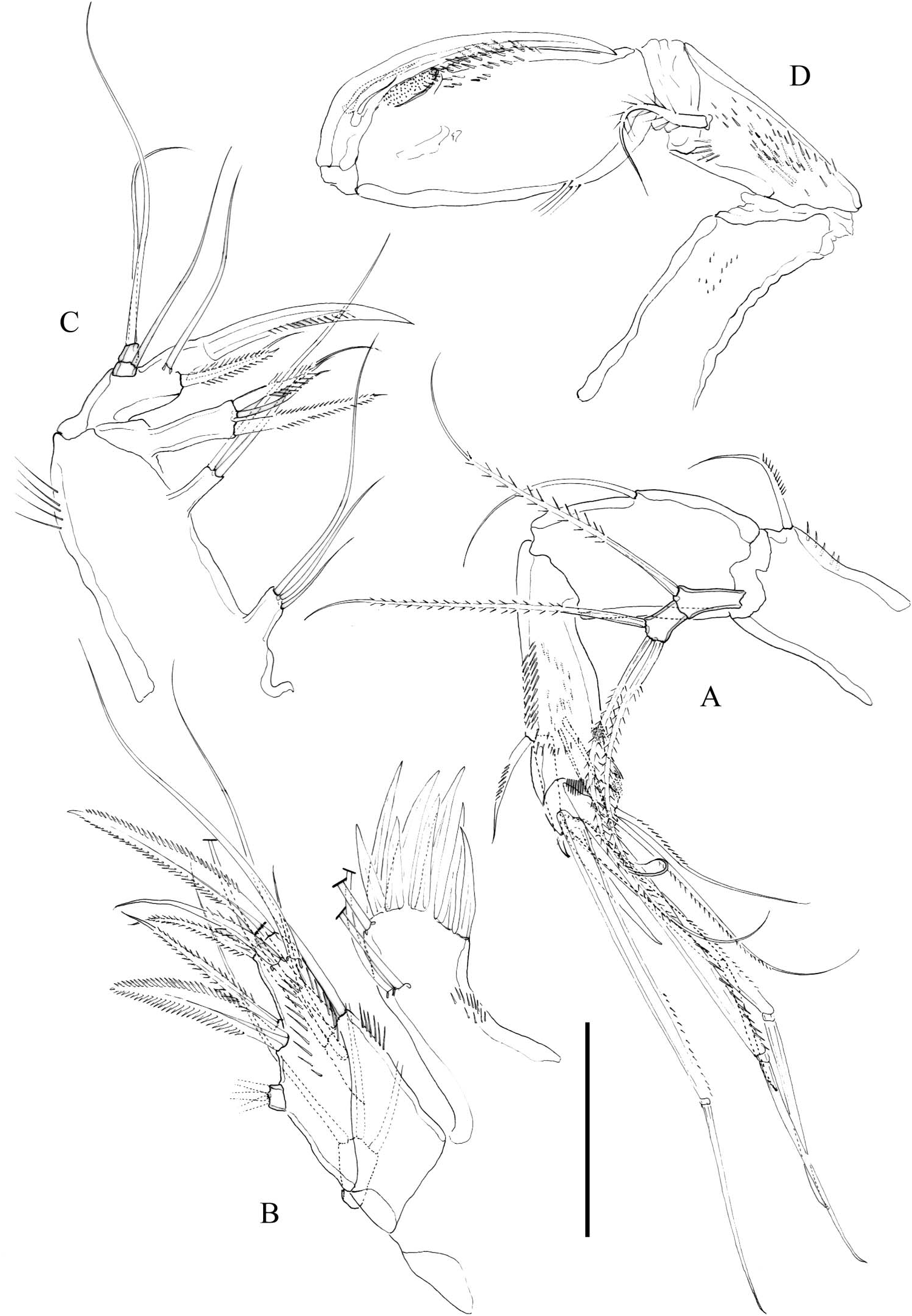
Antenna ( Figure 4A View Figure 4 ). Basis with some inner spinules and one seta. Exopod two segmented; first segment with one, second segment with three setae. Endopod twosegmented; first segment with one inner seta; second segment with small spinules as shown, with two spines and two slender setae laterally, and one spine, four geniculate, and two slender setae apically.
Mandible ( Figure 3B View Figure 3 ). Gnathobasis with three strong teeth and two small spines; with one lateral bipinnate, spine-like element. Coxa-basis large, with four elements. Endopod one-segmented, with one lateral and four apical setae. Exopod one-segmented, visibly shorter than exopod, with three setae.
Maxillule ( Figure 4B View Figure 4 ). Arthrite of praecoxa with eight spines and one seta distally, and two surface setae. Coxal endite with four setae. Basis with six elements. Endopod and exopod one-segmented, with three setae each.
Maxilla ( Figure 4C View Figure 4 ). Syncoxa elongate, with three endites; proximal endite with three, middle endite with two, distal endite with three elements as figured. Allobasis drawn into strong claw with three accessory setae (one of them spine-like). Endopod two segmented; first segment with one, second segment with two slender setae.
Maxilliped ( Figure 4D View Figure 4 ) subchelate. Coxa and basis elongate. Basis with spinules as figured and with one well-developed seta. First endopodal segment rather elongate, with spinules as shown, with cup-shaped structure subdistally; second segment with strong claw bearing two accompanying setae.
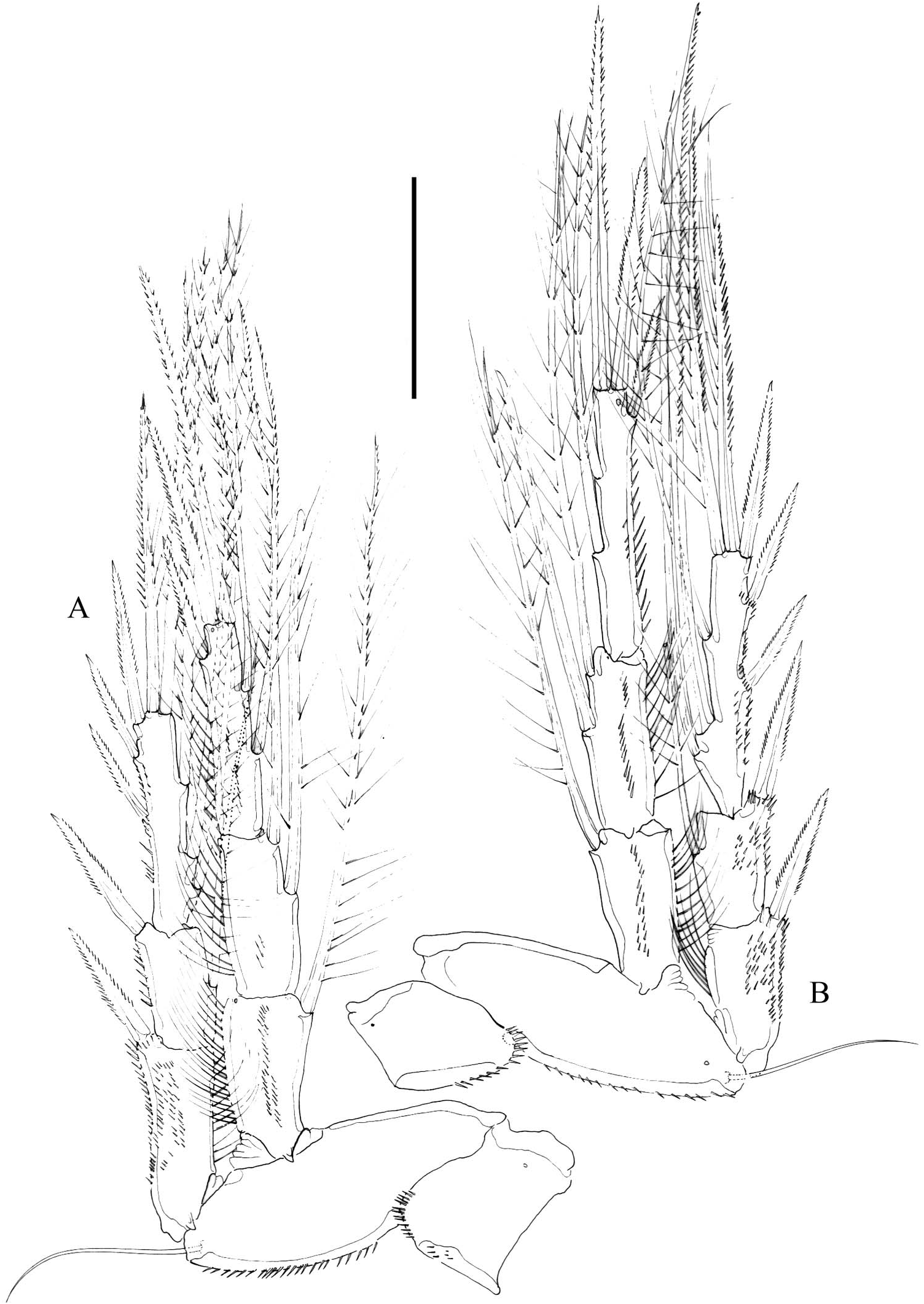
P1 ( Figure 5A View Figure 5 ). Coxa elongate, ornamented as figured. Basis transversally elongate, ornamented with spinules as figured; with slender naked outer seta, and bipinnate inner element. Exopod three-segmented, slightly curved; first segment elongate, about 3.3 times as long as wide, with one outer seta; second segment elongate, slightly longer than preceding segment, with one outer and one inner element; third segment small, with four elements apically (one geniculate seta and three pinnate claws (innermost longest, outermost smallest)), and one outer slender seta. Endopod three-segmented, visibly shorter than EXP1 and EXP2 combined; first segment longest, with one inner seta; second segment slightly shorter than preceding segment, without armature; third segment smallest, with one outer, two apical, and two inner setae.
P2 ( Figure 5B View Figure 5 ). Coxa quadrate. Basis transversely elongate, ornamented as shown, with outer bipinnate seta. Exopod three-segmented; first segment without, second segment with one inner seta; third segment with two inner and two apical elements, and three outer spines. Endopod three-segmented, slightly shorter than exopod; first segment with one, second segment with two inner setae (proximal seta shorter, both setae of about the same thickness); third segment with one outer spine, two apical elements, and two inner setae.
P3 ( Figure 6A View Figure 6 ). Coxa and basis as in P2 except for shorter spinules. Exopod threesegmented, shorter than endopod; first segment without, second segment with one inner seta; third segment with three inner setae subequal in thickness, two apical elements, and three outer spines. Endopod three-segmented, slightly longer than exopod; first segment with one, second segment with two inner setae (proximal seta somewhat stronger); third segment with one outer spine, two apical elements, and two inner setae.
P4 ( Figure 6B View Figure 6 ). Coxa and basis as in P3. Exopod three-segmented, visibly shorter than endopod; first segment without, second segment with one inner seta; third segment with three inner setae (middle seta visibly stronger), two apical elements, and three outer spines. Endopod three-segmented, longer than exopod; first segment with one, second segment with two inner setae (proximal seta shorter and somewhat stronger); third segment with one outer spine, two apical elements, and two inner setae.
P5 ( Figure 2C View Figure 2 ). Baseoendopod elongate, covered with minute spinules as figured; inner baseoendopodal lobe weakly developed, with two spine-like elements distally and three slender inner setae, plus outer slender basal seta. Exopod elongate, about 4.7 times as long as wide, covered with minute spinules; with three apical and three outer elements (most elements lost; their positions arrowed in Figure 2C View Figure 2 ).
Armature formulae as in Table 1.
Male. Body ( Figure 7 View Figure 7 ) as in female, except for separate second (P6-bearing somite) and third urosomites. Total body length of allotype, 725 µm measured from tip of rostrum to posterior margin of caudal rami. Caudal rami ( Figure 8A View Figure 8 ) as in female.
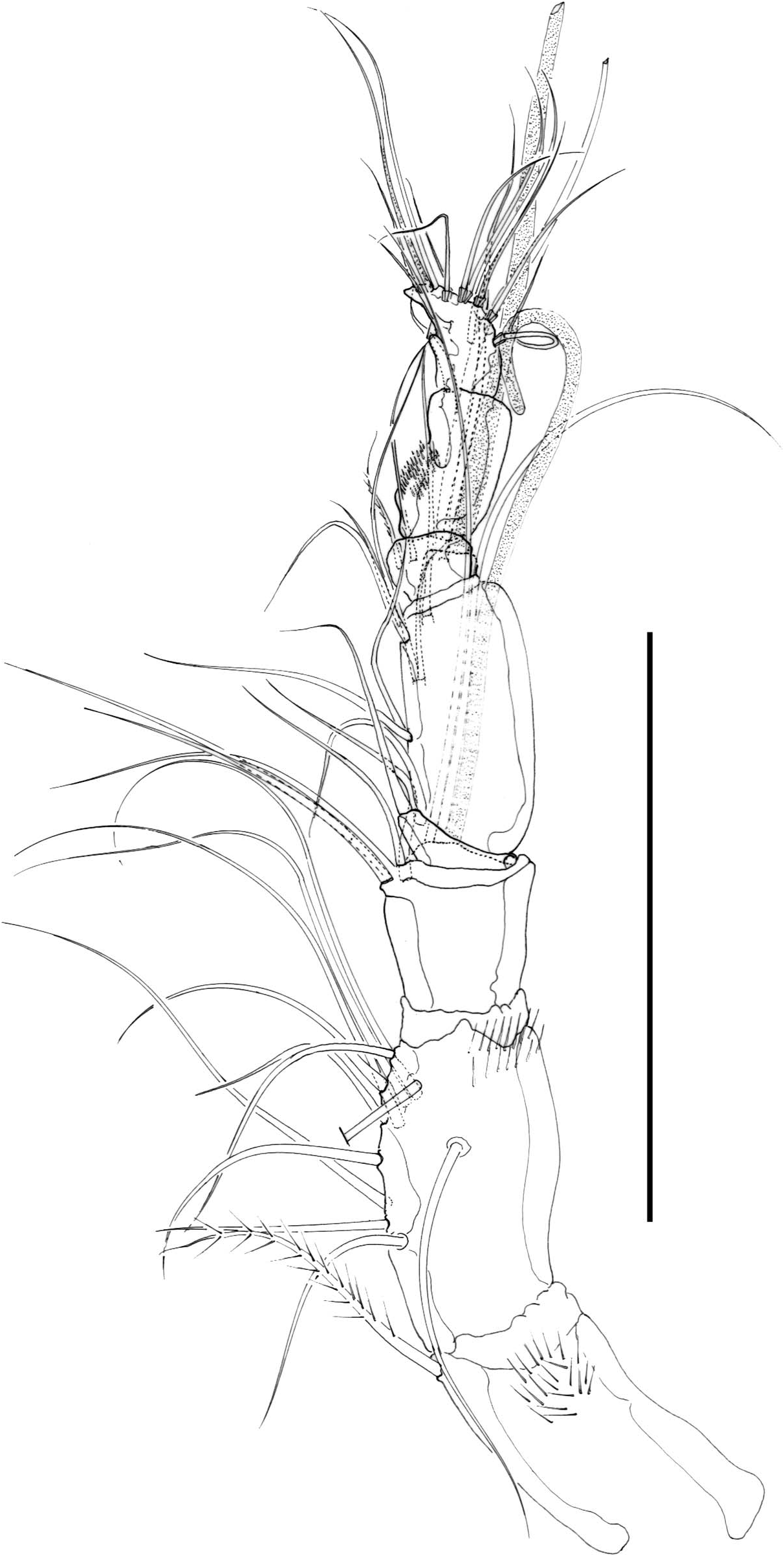
Antennule ( Figure 3B View Figure 3 ) eight-segmented, haplocer. First, second and seventh segments ornamented as figured; with just a few pinnate setae (most setae naked and slender). Armature formula difficult to define, as follows: I – (1); II – (12); III – (7 + ae); IV – (2); V – (9 + ae); VI – (2); VII – (1); VIII – (12 + ae).
Antenna, mandible, maxillule, maxilla and maxilliped as in female.
P1–P4 as in female.
P5 ( Figure 8B View Figure 8 ). Baseoendopod elongate, covered with minute spinules; inner baseoendopodal lobe represented by one element, with outer basal seta. Exopod elongate, covered with minute spinules; with three apical elements and one outer seta.
P6 ( Figure 8A View Figure 8 ). Each leg represented by small lobe with two setae each (outer element smaller).
Remarks
Baird (1837) presented a very brief description of Cyclops depressus (now A. depressa ) in which he illustrated the female and male habitus, the P1 (which Baird described as having one inner seta on P1 ENP1 and ENP2, but the armature of P1 EXP2 is not mentioned in the description, and the figure lacks sufficient detail), and the male P5. Nothing further of the morphology of P2–P4 was described. Later on, Baird (1845) coined the name for the genus Alteutha . Sars (1904) noted that Baird’s (1837) description of A. depressa was very poor for the standards of the epoch, and even though he was not completely certain about the identity of his material, he accepted Brady’s view and concluded that the Norwegian material reported by him might belong to A. depressa ( Baird, 1837) . Interestingly, Sars (1904) did not describe the armature formula of the P2–P4 EXP1, but he did illustrate the P2, showing the P2 EXP1 devoid of inner armature, and the P1 ENP2 without inner seta (contrasting with Baird’s [1837] original description). Also, Sars (1904) and Brady ( Sars 1904) suggested that A. oblonga and A. purpurocincta Norman, 1869 might be assignable to A. depressa , and Monard (1928) regarded the latter species and Claus’ (1863) A. messinensis Claus as distinct. Monard (1928) admitted that the description in Claus (1863) was very poor, and, like Claus (1863), he also remained silent regarding the armature formula of P2– P4 EXP1 of A. messinensis , and described the P1 with one inner seta on ENP2 and one inner seta only on EXP2.
Lang (1948) did not follow Sars and Brady’s view ( Sars 1904) and considered A. purpurocincta and A. depressa sensu Sars (1904) as synonyms of A. oblonga , keeping A. depressa and A. oblonga as distinct species, the former being diagnosed with five setae on A2 EXP (with four setae only in A. oblonga ), with an inner seta on P1 ENP2 (without inner seta in A. oblonga ), and one inner seta only (without outer seta) on EXP2 (the armature formula of P1 EXP2 of A. oblonga remains unknown), the armature formula of P2–P4 EXP1 being unknown. Briefly, Lang (1948) recognized the genus to be composed of A. depressa , A. interrupta (Goodsir, 1845) , A. oblonga , A. novae-zealandiae (Brady, 1899) , A. signata Brady, 1910 , A. austrina , A. dubia T. Scott, 1912 , A. sarsi Monard, 1924 , and A. trisetosa . At present, there are 24 valid species of Alteutha ( Boxshall and Huys 2012) : the above species, except for A. austrina which is currently considered as a synonym of A. depressa ( Hicks 1982) , A. langi Monk , A. polarsternae Dahms , A. rara Jakobi , A. roeae Hicks , A. sewelli Krishnaswamy , A. typica Czerniavski and A. potter Veit-Köhler and Fuentes.
Pallares (1968) described A. littoralis Pallares from Puerto Deseado and Sorrel ( Santa Cruz, Argentina) and presented a synoptic table in which she ( Pallares 1968, p. 252) described the differences observed between her A. littoralis , A. sarsi and A. depressa . Seemingly, Pallares (1968) proposed her A. littoralis as a new species based on the differences she observed. Later, Pallares (1979) found some peltidiids which she attributed to A. depressa , but omitted any comment about the morphology of her specimens and, probably following Lang (1948) and the description of A. depressa from Tristan da Cunha by Wiborg (1964), relegated her A. littoralis to a junior synonym of A. depressa . Note that, contrary to the diagnosis presented by Lang (1948), Pallares (1968) and Wiborg (1964) described the P1 EXP2 with one inner and one outer seta and, surprisingly, Wiborg (1964) described the P2-P4 ENP1 with two inner setae, casting doubts about the identity of Wiborg’s (1964) material. The same applies for A. depressa given the poor, sketchy, and sometimes contradictory descriptions of most records of the species. Pallares’ (1968) view was followed by Hicks (1982) who also synonymized A. langi , A. signata sensu Nicholls (1941) , A. littoralis sensu inter alia Hicks (1977) and A. austrina with A. depressa . Veit-Köhler and Fuentes (2007, p. 157) listed a number of character states shared by all species of Alteutha and noted that only some species lack the inner seta on P2–P4 EXP1 commonly found in most species. According to Veit-Köhler and Fuentes (2007), the species lacking an inner seta of P2–P4 EXP1 are A. trisetosa , A. rara and A. oblonga . The same authors noted also that A. austrina , “synonymized under Alteutha depressa ( Baird, 1837) by Hicks (1982) ” ( Veit-Köhler and Fuentes 2007, p. 157), lacks such seta, implicitly suggesting A. austrina to be different from A. depressa . Probably this decision was based on the material of A. depressa described by Wiborg (1964) from Tristan da Cunha, on Pallares’ (1968, 1979) papers, and in the only male found by Hicks (1982) from Algoa Bay ( South Africa) in which an inner seta is present in P2–P4 EXP1. This decision might also have been supported by the lack of an inner seta in the specimens of A. austrina from Scotia Bay (South Orkneys) upon which Scott (1912) based the description of the species, as well as in the material reported by Pallares (1968) from Santa Cruz ( Argentina).
The material presented herein clearly belongs to the group of species without inner armature in the P2–P4 EXP1 ( A. trisetosa , A. rara , A. oblonga and A. austrina ). Of these species, A. trisetosa and A. austrina possess an inner seta on P1 ENP2. Alteutha rara and A. oblonga lack the inner seta on P1 ENP2, and seem to be more closely related to the Mexican A. alsagopu sp. nov. The morphology of the Mexican specimens matches more closely that of A. oblonga collected off the west coast of Norway and from Trondheim Fjord by Sars (1904), and from Tierra del Fuego ( Argentina) by Pallares (1979), given the armature of the A2 EXP and of P1 EXP2. The former structure is two-segmented, with two and three setae on the first and second segment respectively in A. rara , but two-segmented and with one and three setae on first and second segment respectively in A. oblonga and A. alsagopu sp. nov. The P1 EXP2 bears one inner and one outer seta in A. oblonga and A. alsagopu sp. nov., but with one outer seta only in A. rara . Sars’ (1904) figures of the mouth parts, caudal ramus and swimming legs correspond to the description of the Mexican species and the only differences found between the Mexican species and Sars’ (1904) material of A. oblonga are: (a) the position of the lateral seta of the first endite of the maxilla (displaced laterally in Sars’ [1904] material, but almost apical in the Mexican species); and (b) P1 EXP:ENP length ratio (endopod as long as two first exopodal segments combined in Sars’ [1904] A. oblonga , but noticeably shorter in the Mexican species). Unfortunately, Brady (1878) and Pallares (1979) only presented a written description of their material. Brady’s (1878) description is shallow, and the description of the Mexican species matches better with Pallares’ (1979) description in all respects, but no comparisons could be made regarding the P1 EXP:ENP length ratio, the relative position of the setae / spines of bucal appendages or the swimming legs.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |