
Saltoposuchus connectens Huene 1921
|
publication ID |
https://doi.org/ 10.1093/zoolinnean/zlad035 |
|
DOI |
https://doi.org/10.5281/zenodo.8432530 |
|
persistent identifier |
https://treatment.plazi.org/id/03A2222F-3111-9F0B-42B4-FD833975FBE1 |
|
treatment provided by |
Plazi |
|
scientific name |
Saltoposuchus connectens Huene 1921 |
| status |
|
Saltoposuchus connectens Huene 1921
Diagnosis: A medium-sized non-crocodyliform crocodylomorph (femoral length SMNS 12596: 106 mm) distinguished by the following combination of character states (autapomorphies among non-crocodyliform crocodylomorphs are indicated with an asterix): enlarged anterior maxillary teeth (also present in Junggarsuchus sloani ); antorbital fossa well defined, forming a complete circumference around the antorbital fenestra (also present in CM 29894, Kayentasuchus walkeri , and Dromicosuchus grallator ); antorbital fenestra that is approximately half the size of the orbit*; an oval fossa on the ventral surface of the palatine outlined by a radially striated surface medially*; a dorsally expanded anterior end of the dentary (also present in Trialestes romeri and Hesperosuchus agilis, AMNH FARB 6758); splenial slightly involved in the symphysis (also present in Kayentasuchus walkeri ); and a notch on the lateral side of the proximal head of the tibia*. Saltoposuchus connectens is additionally distinguished from Terrestrisuchus gracilis and Litargosuchus leptorhynchus in possessing a longitudinal crest along the dorsolateral surface of the squamosal; a distinct depression for the supratemporal fossa on the posterior end of the dorsal surface of the frontal; strongly sculpted and transversely wide osteoderms; a wide and short, robust ischial shass; and a robust femur that is longer than the tibia and fibula.
Holotype: SMNS 12597 View Materials a–d ( Fig. 2A–E View Figure 2 ), an associated skeleton comprising a partial skull and much of the postcranium. An isolated less angular SMNS 12597 View Materials e ( Fig. 2F View Figure 2 ) and isolated right calcaneum SMNS 12597 View Materials f ( Fig. 2G, H View Figure 2 ) were previously considered to belong to the holotype specimen ( Huene 1921) but are here shown to not belong to the same individual as the holotype (see descriptions of respective bones below). Their assignment to Saltoposuchus connectens is tentatively maintained based on their corresponding morphology to other elements in SMNS 12597 View Materials a–d.
Referred material: SMNS 12597e–f ( Fig. 2F–H View Figure 2 ), isolated less angular and isolated right calcaneum; SMNS 12596 ( Figs 3–5 View Figure 3 View Figure 4 View Figure 5 ), a disarticulated skeleton including several skull elements; SMNS 55009 ( Figs 6 View Figure 6 , 7 View Figure 7 ), several postcervical vertebrae and part of the right hindlimb, including an incomplete pes; SMNS 12591a ( Figs 8 View Figure 8 , 9 View Figure 9 ), a largely complete, strongly dorsoventrally compressed skull and mandible. This specimen has undergone acidbased preparation since the description of this specimen by Sereno and Wild (1992), revealing additional morphological information, particularly on the palate ( Fig. 9C, D View Figure 9 ); SMNS 12352 ( Fig. 10 View Figure 10 ), a largely complete, well preserved snout, and a less manus.
Locality and horizon: The discontinued ‘Weisser Steinbruch’ sandstone quarries near Pfaffenhofen. Middle Stubensandstein Member of the Löwenstein Formation, middle Keuper (Norian, Late Triassic; Sues and Schoch 2013).
Ontogenetic assessment: The relative sizes of the Saltoposuchus connectens specimens are compared in Table 2. Size discrepancy between the smallest (SMNS 12591a) and largest specimen (SMNS 55009) is inferred at 58.1%. The right femur of SMNS 12596, the second-largest specimen (90.2%), has been histologically sectioned. The interpretation of its relative age is somewhat ambiguous but the individual probably represents a sub-adult, since it is neither an early juvenile, based on the relative size and robusticity of its skeleton, nor skeletally mature, based on the presence of open neurocentral sutures in some of the presacral vertebrae and the absence of an outer circumferential layer (OCL; also known as an external fundamental system, EFS) and secondary osteons in the histological section (see discussion below).
Description
Skull
Premaxilla: SMNS 12352a is the only specimen to preserve remains of the premaxilla and even in this specimen only a badly broken right premaxilla and a small fragment of the posterodorsal process of the less premaxilla are present ( Fig. 10A, B View Figure 10 ). The premaxilla connects to the maxilla posteriorly and to the nasal posterodorsally. The right premaxilla preserves two teeth and is incomplete anteriorly. It bears a prominent posterodorsal process, which wedges between, and overlaps with, both the maxilla and nasal, as in other non-crocodyliform crocodylomorphs like Dromicosuchus grallator ( Sues et al. 2003) , differing from the tight suture between the premaxilla and maxilla seen in crocodyliforms (e.g. Colbert and Mook 1951). Thus, the maxilla did not contribute to the margin of the external naris in Saltoposuchus connectens , as was also indicated by Sereno and Wild (1992).
Maxilla: The maxilla is preserved in SMNS 12596a, SMNS 12352a, and SMNS 12591a, and it thus allows for comparison betweenaspecimenunambiguouslyaưributableto Saltoposuchus connectens (SMNS 12596) and specimens with a controversial taxonomic identification (SMNS 12352, SMNS 12591a; Table 1). In fact, most of the features put forward to distinguish these specimens taxonomically have revolved around the maxilla ( Ostrom 1981, Knoll and Rohrberg 2012). SMNS 12596a preserves a disarticulated, virtually complete, less maxilla ( Fig. 3A View Figure 3 ). Based on the length of this element, this specimen is considerably larger than SMNS 12352 and SMNS 12591a ( Table 2). It preserves four teeth, with the largest, the second counted from anterior, being partially shissed out of its alveolus, exposing much of the root. At least eight nutrient foramina are present dorsal to the alveolar margin in lateral view. SMNS 12352a has a virtually complete right maxilla, which preserves 14 teeth ( Fig. 10A, B View Figure 10 ). A single tooth is clearly missing and therefore the maxillary tooth count is tentatively established at 15. Many nutrient foramina can be observed dorsal to the alveolar margin. Both maxillae of SMNS 12591a preserve nine teeth, an incomplete count ( Fig. 8 View Figure 8 ). On both sides, the posteriormost teeth are only represented by small protuberances. The posterior extent of the maxillary teeth is similar to that in SMNS 12352a, namely roughly at the level of the ventral foot of the lacrimal. This thus corroborates the interpretation of Sereno and Wild (1992) and opposes Ostrom (1981), who considered both specimens to be distinguishable taxonomically in part based on a difference in posterior extent of the maxillary teeth. The angle formed between the main body of the maxilla and its ascending process is 24° in SMNS 12596a, 29° in SMNS 12352a, and 9° in SMNS 12591a. The very sharp angle of SMNS 12591a can clearly be aưributable to the considerable dorsoventral compression of this specimen, as has been pointed out by previous authors (e.g. Sereno and Wild 1992). The alveolar margin is slightly convex in lateral view in its anterior half and very slightly concave in its posterior half in both SMNS 12596a and SMNS 12352a. In SMNS 12591a this curvature is much less distinct, but this too is clearly aưributable to its dorsoventral compression. This compression also led to the misinterpretation of Ostrom (1981) that the antorbital fenestra of SMNS 12591a is restricted to the anterior portion of the antorbital fossa, and that the lacrimal had a considerable extent anteriorly. The element considered to be the anterior part of the lacrimal in the antorbital fossa in fact represents the palatine, which has been displaced dorsally on its medial side due to dorsoventral compression, as was already indicated by Huene (1921: fig. 22), and which has been corroborated by other authors ( Sereno and Wild 1992, Allen 2010).
The outline of the antorbital cavity and the relative contribution of the antorbital fenestra and fossa are virtually identical between SMNS 12596a and SMNS 12352a ( Figs 3A View Figure 3 , 10A, B View Figure 10 ). Both have a considerable anterior contribution of the antorbital fossa, but neither form a pit in the anteroventral corner of the fossa, in contrast to Terrestrisuchus gracilis ( Crush 1980, 1984, contra Clark 1986). The interpretation by Knoll and Rohrberg (2012) that the anterior corner of the antorbital fenestra is more acute in SMNS 12596a, and that in SMNS 12352a the anterior end of the antorbital fenestra diverges from the ventral margin of the antorbital fossa, are both rejected. Both specimens clearly possess a rounded anterior end of the antorbital fenestra and the anterior outline of the antorbital fossa is virtually indistinguishable between both specimens. The relative distribution of the antorbital fossa within the antorbital cavity is less extensive in SMNS 12591a, but this is aưributable to the incomplete preservation of the thin lamina that comprises the antorbital fossa ( Fig. 8 View Figure 8 ). The considerable extent of the thin lamina of the antorbital fossa within the antorbital cavity, and the corresponding relatively small opening of the antorbital fenestra, seen particularly in SMNS 12596a and SMNS 12352a ( Figs 3A View Figure 3 , 10A, B View Figure 10 ), reflects the anatomy seen in most non-crocodyliform crocodylomorphs except Carnufex carolinensis (e.g. Crush 1984, Walker 1990, Ruebenstahl et al. 2022), but conflicts with the morphology seen in early theropod dinosaurs, including coelophysoids ( Raath 1977, Colbert 1989, Nesbiư et al. 2009). The acid preparation of SMNS 12591a has also revealed much of its palatal region, including what are very likely the posterior portions of the palatal processes of the maxillae ( Fig. 9C, D View Figure 9 ). They meet in the midline and are located quite far posteriorly in the snout. This provides a clear indication of the crocodylomorph affinities of this specimen, as it represents a typical character of this group that foreshadows the maxillary ‘secondary palate’ of later crocodylomorphs ( Nesbiư 2011, Dollman and Choiniere 2022).
Nasal: Clearly identifiable nasals are preserved in SMNS 12352a and SMNS 12591a. In addition, Crush (1980) identified a flat bone fragment on SMNS 12597b as a poorly preserved nasal. However, due to the incompleteness of this element it is not possible to corroborate this identification with confidence, although its preserved morphology does correspond with that of the nasal ( Fig. 2C View Figure 2 ). In SMNS 12352a and SMNS 12591a, the nasals are mostly complete, although their anterior ends are missing ( Figs 9A, B View Figure 9 , 10A, B View Figure 10 ). The nasals are longitudinally elongate and transversely narrow elements that articulate with the premaxilla anteriorly, the maxilla and lacrimal laterally, the prefrontal posterolaterally, and the frontal posteriorly. The nasal gradually widens posteriorly and has straight lateral and medial margins. Posteriorly, the nasals form a V-shaped articulation with the frontals, which wedge between the nasals anteromedially, as can best be observed in SMNS 12352a ( Fig. 10A, B View Figure 10 ). This configuration corresponds to that seen in many early crocodylomorphs ( Crush 1984, Colbert 1989, Walker 1990, Ruebenstahl et al. 2022) and several other Triassic pseudosuchians ( Nesbiư 2011: ch. 43). The dorsal surface of the nasals is distinctly piưed in SMNS 12352a ( Fig. 10A View Figure 10 ). In this specimen, the dorsal surface of the nasal is transversely convex, particularly anteriorly, whereas in SMNS 12591a this surface is flat, resulting from its dorsoventral compression.
Lacrimal: Both lacrimals are preserved in both SMNS 12352a and SMNS 12591a, although only a portion of the shass is exposed for the less lacrimal of SMNS 12352a ( Figs 8 View Figure 8 , 9 View Figure 9 , 10A–D View Figure 10 ). Although both lacrimals are virtually complete in SMNS 12591a, they are deformed and poorly preserved, and the right lacrimal of SMNS 12352a is the best representative of this element. It has an inverted L-shape, with a dorsoventrally oriented shass and an anterodorsal process ( Fig. 10A, B View Figure 10 ). At its ventral end, the lacrimal connects to the jugal posteriorly and the maxilla anteriorly. Posterodorsally, the lacrimal connects to the prefrontal, dorsomedially to the nasal, and anteriorly again to the maxilla. The lacrimal also forms the anteroventral portion of the orbital margin and the posterodorsal and posterior margins of the antorbital fenestra. The shass has a prominent anterolateral crest along its ventral half, and the portion of the antorbital fossa directly anterior to it is thus deeply excavated ( Fig. 10A, B View Figure 10 ), similar to the condition seen in crocodylomorphs like Litargosuchus leptorhynchus , Dromicosuchus grallator , Carnufex carolinensis , and Dibothrosuchus elaphros ( Clark and Sues 2002, Sues et al. 2003, Drymala and Zanno 2016, Ruebenstahl et al. 2022). An anteriorly directed opening within a concavity on the posterior margin of the lacrimal shass near the tapering ventral end of the prefrontal probably represents the opening for the lacrimal duct, as was also indicated by Sereno and Wild (1992), although this cannot be determined with certainty. The ventral foot of the shass extends posteroventrally to form a confluent connection with the jugal, similar to that of all other non-crocodyliform crocodylomorphs except Carnufex carolinensis , which lacks a subcircular orbit ( Drymala and Zanno 2016).
The lacrimal forms a large contribution to the antorbital fossa both directly anterior to the lacrimal shass and along the anterodorsal process ( Fig. 10A, B View Figure 10 ). The former contribution represents an anteroposteriorly narrow lamina, whereas the laưer contribution is considerable, entirely filling the dorsal half of the antorbital cavity at its greatest extent, near its posterior end. The antorbital fossa contribution of the anterodorsal process gradually reduces in height anteriorly, providing the process with a triangular outline in lateral view. This condition is similar to that seen in most non-crocodyliform crocodylomorphs, including Terrestrisuchus gracilis , Dromicosuchus grallator, CM 29894, and Carnufex carolinensis (e.g. Crush 1984, Clark and Sues 2002, Sues et al. 2003, Drymala and Zanno 2016), except for Sphenosuchus acutus and Junggarsuchus sloani , in which the antorbital fossa contribution of the anterodorsal process of the lacrimal is much more restricted ( Walker 1990, Ruebenstahl et al. 2022). Note that CM 29894 was originally referred to Hesperosuchus agilis ( Clark et al. 2000) . Since then, it has been suggested that this specimen, as well as an additional crocodylomorph specimen from the same locality, YPM 41198, might be referable to a separate taxon, based in part on an approximately 20 million year gap between these two specimens, which come from the Coelophysis ºuarry in the upper part of the Chinle Formation, and the holotype of Hesperosuchus agilis, AMNH FARB 6758, which derives from the Blue Mesa Member in the lower part of the Chinle Formation ( Nesbiư 2011, Irmis et al. 2013). Although others have suggested that there are no distinct morphological distinctions between AMNH FARB 6758 and CM 29894 ( Leardi et al. 2017), to avoid confusion separate morphological comparisons are made here to CM 29894 and YPM 41198, and to Hesperosuchus agilis as represented exclusively by the holotype AMNH FARB 6758. The exact anterior extent of the lacrimal and its contribution to the antorbital fossa is hard to establish in SMNS 12352a, but it appears to reach to close to the anterior end of the antorbital fenestra. In both lacrimals of SMNS 12591a, the contribution to the antorbital fossa is almost completely missing.
Preffontal: The prefrontal is also preserved in SMNS 12352a and SMNS 12591a. The right prefrontal of SMNS 12352a is well preserved but incomplete, only preserving its anteriormost and ventral portions ( Fig. 10A, B View Figure 10 ). Both prefrontals of SMNS 12591a are virtually complete, but their anterior and ventral portions are poorly preserved due to compression ( Figs 8 View Figure 8 , 9 View Figure 9 ). The prefrontal forms most of the anterior margin of the orbit and articulates with the lacrimal anteriorly and the nasal and frontal dorsomedially. In SMNS 12591a, the prefrontal also connects extensively with the palate, and although this is also the case in several other non-crocodyliform crocodylomorphs (e.g. Nesbiư 2011, Ruebenstahl et al. 2022), it cannot be excluded that this is the result of the dorsoventral compression in this specimen. The ventral process of the prefrontal forms a thin strip of bone that tapers ventrally and reaches until a level just ventral to the dorsoventral midpoint of the orbit in lateral view. In dorsal view, the prefrontal has a markedly convex lateral margin, which is best observed on the right side of SMNS 12591a. A similar convexity is also present in Junggarsuchus sloani , which was described as a prefrontal overhang of the orbit in that taxon ( Ruebenstahl et al. 2022). The medial margin is more difficult to discern due to poor preservation, but it is roughly straight in dorsal view. Posteriorly, the prefrontal reaches until just anterior to the dorsal apex of the orbit. As was also indicated by Sereno and Wild (1992), the prefrontal extends similarly far posteriorly on the ventral side of the skull roof. The presence of a considerable medial extension of the prefrontal on the anterior side of the orbit is widely present among crocodylomorphs ( Nesbiư 2011: ch. 40) and this feature can also be observed on the right prefrontal of SMNS 12591a.
Frontal: Disarticulated but virtually complete frontals are preserved and exposed in ventral view in SMNS 12597a ( Figs 2A View Figure 2 , 11A View Figure 11 ). Both frontals are also virtually complete in SMNS 12591a and the anterior ends of the frontals are preserved in SMNS 12352a ( Figs 9A, B View Figure 9 , 10A, B View Figure 10 ). The frontal is an elongate element with a straight medial margin, which articulates with its antimere, and a more complex lateral margin that for most of its length is distinctly concave in dorsal and ventral view. This concavity forms the dorsal margin of the orbit. At its anterior end, the frontal mostly articulates with the nasal, but also with the prefrontal on its lateral side. As outlined above, the articulation between frontals and nasals is V-shaped, with each frontal possessing a distinctly medially tapering anterior end. The frontalnasal region is badly broken in SMNS 12591a, but the V-shaped suture between the elements can still be discerned. The frontal articulates with the postorbital on its posterolateral end, and with the parietal along its posterior margin. This last articulation is W-shaped in dorsal view in SMNS 12597a and SMNS 12591a, similar to that seen in Terrestrisuchus gracilis , Dromicosuchus grallator , Dibothrosuchus elaphros , and Almadasuchus figarii ( Crush 1984, Sues et al. 2003, Leardi et al. 2020, Ruebenstahl et al. 2022). The frontal is transversely widest near its posterior end, where it articulates laterally with the dorsomedial process of the postorbital.
The dorsal surface of the frontals is mostly flat, but a shallow, straight, longitudinal gulley is present directly medial to the orbital margin and, consequently, the surface directly medial to this gulley is convex. A similar morphology is also seen in Terrestrisuchus gracilis , Litargosuchus leptorhynchus , and Dromicosuchus grallator (NHMUK PV R 37731, Clark and Sues 2002, Sues et al. 2003), whereas a more distinct, medial longitudinal bulge or ridge is present in Hesperosuchus agilis (AMNH FARB 6758) and non-crocodyliform crocodylomorphs that are generally found to be more closely related to crocodyliforms ( Colbert 1952, Nesbiư 2011: ch. 42). The posterolateral end of the less frontal in SMNS 12591a bears a clear concavity on its dorsal surface, which is anteriorly rounded. This concavity was also indicated by Sereno and Wild (1992), and represents the contribution of the frontal to the supratemporal fossa. This frontal contribution is also present in other non-crocodyliform crocodylomorphs (e.g. Colbert 1952, Walker 1990, Ruebenstahl et al. 2022), except for Terrestrisuchus gracilis and Litargosuchus leptorhynchus ( Crush 1984, Clark and Sues 2002).
The ventral surface of the frontals of SMNS 12597a is characterized by a distinct longitudinal ridge, which runs near and parallel to the orbital rim of the bone. The medial edge of the bone is also raised ventrally, and between both ridges the excavated olfactory tract is formed ( Fig 11A View Figure 11 ). Within this tract, a subcircular concavity just anterior to the transversely narrowest point of the frontal forms the olfactory bulb ( Walker 1990). A distinct excavation on the anterolateral margin of the ventral surface of the frontal forms the facet for the prefrontal. The anterolateral facet for the prefrontal and the orbital rim are separated by a triangular process that is directed somewhat ventrally. A similarly shaped process is also present in Terrestrisuchus gracilis and Sphenosuchus acutus (NHMUK PV R 38290b, Walker 1990). On the posterolateral margin of this process, a small but distinct groove is present, which forms an additional articulation facet for the prefrontal. The ventral surface of the posterior end on the right frontal forms a clear facet for the parietal, which thus slightly overlapped the frontal ventrally here. As such, the articulation between the frontal and parietal was complex, with the frontal overlying the parietal dorsally, except for a medial peg on the frontal that articulated on the ventral side of the parietal, as was also described by Crush (1980). The wide posterolateral end of the frontal forms a thin articulation facet for the postorbital, which thus articulated on the ventral side of this frontal process. The posterolateral end of the frontal tapers and terminates in a sharp tip.
Parietal: A partial parietal is preserved in SMNS 12597b. This element was originally identified as a quadratojugal by Huene (1921), but later correctly reinterpreted as the posterior half of a less parietal exposed in dorsal view by Crush (1980), including the posterolateral process ( Fig. 2C View Figure 2 ). The anterior halves of both parietals are also present in SMNS 12591a ( Fig. 9A, B View Figure 9 ). The parietals are clearly unfused in both specimens. Anteriorly, the parietal only connects to the frontal. As outlined above, the frontal–parietal suture is W-shaped in dorsal view, with a distinct anteromedial process of the parietal and a V-shaped notch directly lateral to this process. The dorsal surface of the parietal is smooth, but possesses a marked depression on its lateral half, and correspondingly, a raised longitudinally-directed platform on its medial half. A distinct sagiưal crest, which is present in several non-crocodyliform crocodylmorphs, including Pseudhesperosuchus jachaleri and Sphenosuchus acutus , is absent as in Terrestrisuchus gracilis , Litargosuchus leptorhynchus, CM 29894, and Dromicosuchus grallator ( Crush 1984, Clark et al. 2000, Clark and Sues 2002, Sues et al. 2003). The lateral depression of the parietal is part of the supratemporal fossa, and the lateral margin of the bone, which is concave in dorsal view, forms the medial margin of the supratemporal fenestra. The medial margin, which forms the suture between the parietals, is straight. The posterior margin of the less parietal of SMNS 12597b is straight and slightly raised. It is directed posterolaterally to anteromedially. The occipital surface of the parietal is not exposed.
Postorbital: The postorbital is only preserved in SMNS 12591a ( Figs 8 View Figure 8 , 9A, B View Figure 9 ). It connects to the frontal dorsally, to the jugal ventrally, and it forms an extensive connection with the squamosal along its posterior side. It is a triradiate bone composed of a dorsomedial process, ventral process, and posterior process. The first two processes are elongate, with the ventral process being the longest, whereas the posterior process is considerably shorter and triangular in outline. The dorsomedial and posterior processes are primarily exposed in dorsal view, whereas the ventral process is deflected relative to the rest of the element, and exposed in lateral view. Although this deflection is at least to some extent the result of the dorsoventral compression of SMNS 12591a, the configuration is similar to that seen in Terrestrisuchus gracilis and Litargosuchus leptorhynchus ( Crush 1984, Clark and Sues 2002). The anterior margin of the postorbital is distinctly concave in lateral and dorsal view, and forms much of the posterior margin of the orbit. The connection with the frontal is best preserved on the right side of SMNS 12591a, which reveals that the medial margin of the dorsomedial process articulated tightly with the lateral surface of the posterolateral side of the frontal, thus corroborating the interpretation of the postorbital facet on the disarticulated frontals of SMNS 12597a described above. This corresponds to the morphology seen in early crocodylomorphs, whereas in later non-crocodyliform crocodylomorphs, such as Dibothrosuchus elaphros and Junggarsuchus sloani , the dorsomedial process additionally articulates with the parietal ( Ruebenstahl et al. 2022).
The dorsomedial process is distinctly curved anterodorsally. The posterior process, as well as the proximal bases of the dorsomedial and ventral processes, all overlapped the squamosal dorsally. This wide overlapping articulation between postorbital and squamosal does not occur in theropods, and is typical of non-crocodyliform crocodylomorphs ( Sereno and Wild 1992, Nesbiư 2011: ch. 66). The configuration of SMNS 12591a is particularly reminiscent of that seen in Terrestrisuchus gracilis and Litargosuchus leptorhynchus ( Crush 1984, Clark and Sues 2002) and thus represents a clear indication of the crocodylomorph identity of this specimen, as was also pointed out by Sereno and Wild (1992). The ventral process of the postorbital articulates on the anterior side of the ascending process of the jugal and tapers to a point ventrally, contrasting with the condition in Terrestrisuchus gracilis , Litargosuchus leptorhynchus , and Sphenosuchus acutus , in which the tip of the ventral process is gently rounded ( Crush 1984, Walker 1990, Clark and Sues 2002).
Jugal: A largely complete less jugal missing the anterior end of the anterior process is preserved in SMNS 12597b ( Fig. 2C View Figure 2 ). The lateral surface of this element is generally well preserved, but broken in the region where the three processes of the bone meet. This element was originally interpreted as a lacrimal ( Huene 1921), but reinterpreted as a jugal by Crush (1980). The anterior tip of the right jugal is present in SMNS 12352a, whereas both jugals are largely complete in SMNS 12591a ( Figs 8 View Figure 8 , 9 View Figure 9 , 10A, B View Figure 10 ). In the laưer specimen, the edges of the anterior process are broken in the less element, and the distal end of the posterior process is missing in the right element. The jugal is a triradiate element, consisting of an anterior, posterior, and dorsal or ascending process. The anterior and posterior processes are both oriented in the horizontal plane relative to the skull, whereas the ascending process is oriented virtually completely perpendicular to the other two processes in both SMNS 12597b and SMNS 12591a, being directed dorsally and very slightly posteriorly.This contrasts with Terrestrisuchus gracilis and Carnufex carolinensis , in which the ascending process is more posterodorsally oriented (NHMUK PV R 7591, Crush 1984, Drymala and Zanno 2016), but corresponds with the morphology seen in Litargosuchus leptorhynchus and Sphenosuchus acutus ( Walker 1990, Clark and Sues 2002). The anterior process forms the longest of the three processes, followed by the posterior process. The anterior process forms the ventral margin of the orbit, the posterior process forms the ventral margin of the infratemporal fenestra, and the ascending process forms the posteroventral margin of the orbit on its anterior side and the anteroventral margin of the infratemporal fenestra on its posterior side.
The anterior process does not contribute to the antorbital fossa in both SMNS 12352a and SMNS 12591a, similar to other non-crocodyliform crocodylomorphs ( Sereno and Wild 1992, Nesbiư 2011: ch. 69). The anterior articulation of the jugal with the maxilla and lacrimal is best observed in SMNS 12352a. The jugal sits in a slot on the dorsolateral surface of the maxilla, whereas the jugal connects to the lacrimal on the dorsomedial surface of the former and the ventral surface of the laưer. The ascending process has a straight to slightly convex posterior margin in lateral view in SMNS 12597b and both jugals of SMNS 12591a, a character unique to certain early crocodylomorphs among archosaurs ( Nesbiư 2011: ch. 73), including Sphenosuchus acutus , Litargosuchus leptorhynchus , and Junggarsuchus sloani ( Walker 1990, Clark and Sues 2002, Ruebenstahl et al. 2022). Dorsally, the postorbital articulates on the anterior surface of the ascending process, and a faint ridge on the lateral surface of the process demarcates this articular facet on the jugal of SMNS 12597b. At its distal end, the posterior process would have articulated with the quadratojugal on the medial surface of the former, but this articulation is not clearly preserved in any of the specimens. The posterior process gradually tapers posteriorly in lateral view, as can be observed in SMNS 12597b and the less jugal of SMNS 12591a, and terminates in a slightly rounded tip in SMNS 12597b. The lateral surface of the jugal is smooth and transversely convex for most of its length.
Squamosal: Both squamosals are preserved in SMNS 12597b ( Fig. 2C View Figure 2 ). Crush (1980) considered the less squamosal to be beưer preserved but this element has since been damaged, particularly near its posterior end. Thus, the right element is now the more complete and beưer preserved of the two ( Fig. 11C View Figure 11 ). SMNS 12596a possesses a largely complete less squamosal and the anterior portion of a right squamosal is preserved in SMNS 12591a ( Figs 3A View Figure 3 , 8A, B View Figure 8 , 9A, B View Figure 9 , 11B View Figure 11 ). Huene (1921) originally interpreted the squamosals in both SMNS 12597b and SMNS 12596a as pterygoids, but their identification as squamosals was later established by Crush (1980). Allen (2010) misidentified the right squamosal of SMNS 12591a as a displaced quadrate, but its unambiguous identity as a squamosal has been established by Huene (1921) and Sereno and Wild (1992). The squamosal connects to the postorbital anteriorly, the quadrate and quadratojugal ventrally, the parietal posteromedially, and the otoccipital posteroventrally. Its medial edge forms the lateral and posterolateral margin of the supratemporal fenestra, whereas its lateroventral edge forms the dorsal margin of the infratemporal fenestra.
The squamosal is predominantly made up of a large plate-like surface that is curved strongly medially near its posterior end in dorsal view. This plate is transversely wide in all specimens, which represents a characteristic trait of several non-crocodyliform crocodylomorphs, including Terrestrisuchus gracilis , Litargosuchus leptorhynchus , Junggarsuchus sloani , and Almadasuchus figarii (e.g. Crush 1984, Clark and Sues 2002, Ruebenstahl et al. 2022). In SMNS 12591a, the squamosal is considerably wider than the supratemporal fenestra in dorsal view ( Sereno and Wild 1992). The plate is transversely distinctly convex on its dorsolateral surface and conversely concave on its ventromedial surface. The dorsolateral surface is mostly smooth but possesses a distinct longitudinal crest along most of its length, which is predominantly oriented anteroposteriorly but which curves distinctly medially near the posterior end of the bone. The dorsal surface of the squamosal of SMNS 12591a is too broken and incomplete to establish the presence of this crest. A similar crest is present in other non-crocodyliform crocodylomorphs, except Terrestrisuchus gracilis and Litargosuchus leptorhynchus ( Crush 1984, Clark and Sues 2002). It represents the demarcation of the contribution of the squamosal to the supratemporal fossa and thus the dorsolateral extent of the jaw adductor musculature, which is placed medial to the crest ( Sereno and Wild 1992). This medial surface would have predominantly faced dorsally, and the surface lateral to the crest predominantly laterally, with the former surface being most prominent. The outer surface of the squamosal also faces predominantly dorsally in Terrestrisuchus gracilis and Litargosuchus leptorhynchus ( Crush 1984, Clark and Sues 2002). The medial margin of the squamosal drops steeply ventrally, as can best be seen in SMNS 12596a, similar to the condition in Terrestrisuchus gracilis (NHMUK PV R 7591). The anterior portion of the dorsolateral surface of the squamosal of SMNS 12596a is incomplete, but a very large articular surface for the postorbital can nevertheless be observed.
Three processes extend from the posterior end of the less squamosal of SMNS 12596a and the right squamosal of SMNS 12597b ( Fig. 11B, C View Figure 11 ). Despite the disarticulation of these elements, the nature of these processes can be established based on observations on Litargosuchus leptorhynchus (BP/I/5237, unpublished CT data) and Terrestrisuchus gracilis (NHMUK PV R 7591), which both have a similar squamosal morphology. The first process extends from the posteromedial end of the squamosal and is dubbed the posteromedial process. It is hook-shaped, and would have formed the articulation with the posterolateral process of the parietal. The second process is positioned directly ventrolaterally to the posteromedial process, and both are separated by a rounded opening, the post-temporal fenestra, whichwassmallandsubcircularin Saltoposuchusconnectens. This second process is named the post-temporal flange and formed the lateral margin of the fenestra. It is projected ventrally and slightly posteriorly at its base, but distally it curves distinctly medially. It is incomplete in both SMNS 12596a and SMNS 12597b but would probably have tapered to a point distally, as in Terrestrisuchus gracilis and Litargosuchus leptorhynchus . The final process represents the largest of the three. It is positioned lateral to the post-temporal flange and is directed posteroventrally.Since it gradually tapers distally in dorsal view, it is roughly triangular in outline and has a flat, posterodorsally directed surface. This represents the articular surface for the paroccipital process of the otoccipital, and this process is therefore named the otoccipital flange. It is thickened on its lateral side but very dorsoventrally thin on its medial side. The ventral surface in SMNS 12597b possesses a clear concavity at the base of the posteromedial process, which represents the articular surface for the dorsal head of the quadrate. Thus, as in other crocodylomorphs, the dorsal head of the quadrate was positioned medially relative to the ventral end of the quadrate.
Quadrate: A partial less quadrate is preserved in both SMNS 12597b and SMNS 12596a ( Figs 2C View Figure 2 , 3A View Figure 3 , 11D, E View Figure 11 ). The former is exposed in posterolateral view and was originally interpreted as a right quadrate by Huene (1921), whereas the laưer is exposed in anterior view and was originally interpreted as a right palatine. The interpretation offered here corroborates that suggested by Crush (1980). SMNS 12591a also preserves distorted remains of both quadrates ( Fig. 8 View Figure 8 ). Only the less quadrate was identified by Sereno and Wild (1992), probably because the right quadrate, which is distinctly displaced medially and anteriorly, was still embedded in matrix at that time. The less quadrate is heavily distorted, whereas the dorsal portion of the right quadrate is in comparatively good condition.
The shass of the quadrate has a mildly but continuously concave posterior margin. The posterolateral surface of the shass is characterized by a distinct and deep excavation in SMNS 12597b and the right quadrate of SMNS 12591a. This excavation fully pierces the shass in SMNS 12597b, but this is almost certainly the result of the shass being partially broken. In SMNS 12597b, the excavation is oval in posterior view, being dorsoventrally taller than mediolaterally wide ( Fig. 11D View Figure 11 ). This excavation represents the internal quadrate fenestra, a pneumatic feature of the crocodylomorph quadrate, which is connected to a system of internal diverticula in hallopodids ( Leardi et al. 2017, 2020). Both in the shape of the fenestra and the lack of internal quadrate diverticula, the morphology of SMNS 12597b is very similar to Terrestrisuchus gracilis and Dromicosuchus grallator (NHMUK PV R 7591, Sues et al. 2003). The remainder of the posterolateral surface of the quadrate is roughly flat to slightly concave. The posterior margin of the quadrate curves strongly posterodorsally towards its dorsal head, which is transversely narrow.
In lateral view, the anterodorsal portion of the quadrate is expanded and terminates in a roughly rectangular end that is confluent with the more posteromedially placed dorsal head in both SMNS 12597b and the right quadrate of SMNS 12591a. This expansion corresponds to the anterodorsal process described by Walker (1990) for Sphenosuchus acutus , although it is considerably anteroposteriorly shorter in Saltoposuchus connectens . Its ventral end of the quadrate is transversely expanded compared to the shass. Its ventral margin is slightly convex in posterior or anterior view. The anterior surface, exposed in SMNS 12596a ( Fig. 11E View Figure 11 ), is mostly transversely concave. The pterygoid ramus is directed anteriorly, medially, and ventrally. Although it is incomplete in SMNS 12596a, it is clear that it is mediolaterally thin and that it gradually reduces in dorsoventral height distally. Its ventral margin is strongly concave proximally, whereas its broken dorsal margin is slightly convex. The ventral margin is thickened relative to the rest of the ramus and its margin is distinctly curved medially. A small ventral projection is present near the distal end of the ramus ( Fig. 11E View Figure 11 ), which might be homologous with the ‘pterygoid process’ of Sphenosuchus acutus ( Walker 1990) . A pterygoid process is also present in the hallopodids Macelognathus vagans and Almadasuchus figarii ( Leardi et al. 2017, 2020).
Quadratojugal: Poorly preserved remains are tentatively identified as the remnants of the less quadratojugal in SMNS 12591a. This is an anteroposteriorly thin bone that is strongly curved so that its anterior margin is strongly convex, and its posterior margin concave, in lateral view ( Fig. 8C, D View Figure 8 ). However, this is probably the result of the distortion if this indeed represents the quadratojugal, as this strongly deviates from the straighter quadratojugals observed in other crocodylomorphs (e.g. Walker 1990).
Vomer: The additional preparation of SMNS 12591a allows for a more extensive description of the palate than was previously possible ( Ostrom 1981, Sereno and Wild 1992), including the first observations of the vomers, which are partially exposed in ventral view and badly cracked ( Fig. 9C, D View Figure 9 ). They are apparently unfused and form elongate, transversely thin, rod-like structures that connect to the palatine and the anterior end of the pterygoid posteriorly and to the palatal contribution of the maxilla anteriorly. Because the vomers are so narrow, the choanae, of which they form the medial margin, are correspondingly wide, similar to the condition seen in Sphenosuchus acutus ( Walker 1990) . In contrast, Gauthier (1986) considered the vomers of SMNS 12591a to be fused anteriorly, which supported a theropod interpretation of this specimen. It is unclear on which this interpretation was based, since it predates the additional preparation that revealed the vomers.
Palatine: Both palatines are present but badly preserved in SMNS 12591a ( Fig. 9C, D View Figure 9 ). The palatine articulates with the pterygoid along its medial margin for most of its length. Additionally, it also connects to the maxilla, and possibly the prefrontal and lacrimal, laterally and the vomer anteromedially. As indicated above, together with the rest of the anterior palatal region, the palatines have been diagenetically compressed dorsally against the nasals and, consequently, they are visible through the antorbital fenestra in lateral view on both sides. The less palatine is best preserved; it is a plate-like bone with a distinctly concave anterior margin in ventral view, which forms the posterior margin of the choana. A distinct rim on the choanal margin, as described for Sphenosuchus acutus , Kayentasuchus walkeri , Dibothrosuchus elaphros , and Junggarsuchus sloani ( Walker 1990, Nesbiư 2011: ch. 91, Ruebenstahl et al. 2022), is absent, as it is in Terrestrisuchus gracilis (NHMUK PV R 38291, Crush 1984). The exact posterior extent of the palatine is unknown but it is clear that the posterior portion is considerably longer than the anterior portion, as is also the case in other non-crocodyliform crocodylomorphs (e.g. Crush 1984, Walker 1990, Clark and Sues 2002, Ruebenstahl et al. 2022). The posterolateral margin is probably incomplete on both sides. However, the preserved portion of this margin, which forms the anterior and anteromedial margins of the suborbital fenestra, is continuously and widely concave in ventral view. The nature of the lateral suture with the maxilla is unclear, while the medial suture with the pterygoid is straight. The ventral surface of the less palatine possesses a very large, oval fossa, which is outlined by a radially striated surface medial to it ( Fig. 9C, D View Figure 9 ). This morphology is unknown for other crocodylomorphs and appears to be unique to Saltoposuchus connectens . The dorsal surface of the palatine as seen through the antorbital fenestra is thickened but otherwise too incompletely preserved to comment on ( Fig. 9A, B View Figure 9 ).
Ectopterygoid: Both ectopterygoids are present but incomplete and poorly preserved in SMNS 12591a, with the right element being best preserved ( Fig. 9 View Figure 9 ). The ectopterygoid contacts the pterygoid posteromedially and the jugal laterally. The ectopterygoid is characterized by a distinctly curved shass that possesses a convex anterior and concave posterior margin in dorsal or ventral view. Furthermore, the lateral side of the ectopterygoid is positioned considerably further dorsally than its posteromedial side. The articular facet with the jugal is composed of an anterior and posterior process, and as such, it is considerably anteroposteriorly longer than dorsoventrally tall, as is also the case in other non-crocodyliform crocodylomorphs (e.g. Walker 1990, Ruebenstahl et al. 2022). The anterior process is displaced ventrally relative to rest of the lateral end, whereas the posterior process curves posterodorsally. The exact articulation with the pterygoid is somewhat unclear, but it is obvious that the ectopterygoid expands lateromedially near its posteromedial end and it articulates on the anterolateral surface of the transverse flange of the pterygoid.
Pterygoid: Both pterygoids are present but incomplete and somewhat poorly preserved in SMNS 12591a ( Fig. 9C, D View Figure 9 ). The anterior portion of the pterygoids is deformed dorsally and crushed against the dorsal roof of the snout. The pterygoid articulates anteriorly with the vomer and anterolaterally with the palatine. Laterally it articulates with the ectopterygoid; the posterolateral articulation with the quadrate is not preserved or exposed. Posterodorsally, the pterygoid articulates with the parabasisphenoid. The palatal ramus of the pterygoid is very elongate and reached approximately until the anterior end of the antorbital fenestra. Its lateral margin gradually tapers anteriorly and is partially ventrally overlapped by the palatine. The medial margin is characterized by a thickened, ventrally directed rim along its entire length, as is also the case in Terrestrisuchus gracilis, CM 29894, Sphenosuchus acutus , and Dibothrosuchus elaphros (e.g. Crush 1984, Walker 1990, Clark et al. 2000, Ruebenstahl et al. 2022). This rim is mostly transversely thin, but it thickens approximately anterior to the level of the palatine–maxilla articulation. Such a transverse thickening is also observed in Sphenosuchus acutus ( Walker 1990) . This thickened anterior portion of the medial rim was interpreted as the vomer by Ostrom (1981), but later correctly identified by Sereno and Wild (1992). The remainder of the ventral surface of the pterygoid is badly broken on both sides but appears to be smooth and mostly flat.
The pterygoid connects to its antimere along the entire length of the palatal ramus and an interpterygoid vacuity was therefore absent. This corresponds to the condition in all other non-crocodyliform crocodylomorphs known from articulated palates with well preserved pterygoids ( Bonaparte 1972, Walker 1990, Clark et al. 2000, Ruebenstahl et al. 2022). An interpterygoid vacuity was considered to be present in Terrestrisuchus gracilis ( Crush 1980, 1984), but this was based on disarticulated elements and this interpretation is therefore uncertain, as was also discussed by Clark et al. (2000). The shape of the transverse flange of the pterygoid is hard to ascertain but it appears to be elongated posterolaterally, as in other non-crocodyliform crocodylomorphs (e.g. Crush 1984, Walker 1990, Ruebenstahl et al. 2022). Posterior to the palatal ramus, the ventral rim of the pterygoid curves posterolaterally where it forms a process that is slightly posteroventrally directed, the medial pterygoid flange (sensu Walker 1990). Its posterior end is covered by the ceratobranchial I ventrally on both sides. A thin, transversely concave lamina is present on the ventral surface medial to this flange in the right pterygoid, and its posteromedial corner clearly articulates with the basipterygoid process of the parabasisphenoid. A short portion of the posterolaterally directed quadrate flange of the right pterygoid can be observed in ventral view, but its morphology is unclear.
Braincase: Incomplete braincase remains are certainly preserved in SMNS 12591a. Because the skull is abruptly broken posteriorly and the elements near this break are severely distorted, most bones cannot be identified. The only braincase element to exhibit sufficient preservation is the parabasisphenoid, which is partially exposed in ventral view ( Fig. 9C, D View Figure 9 ). It was identified by Huene (1921) and also briefly described by Sereno and Wild (1992). The exposed portion of the parabasisphenoid represents the ventral surface, including remains of both basipterygoid processes, which are incomplete. The ventral surface is inclined slightly posterodorsally to anteroventrally relative to the main axis of the skull. Its surface is transversely concave along its length, forming a shallow longitudinal trough that represents the anterior portion of the median pharyngeal or sub-basisphenoid recess. The posterior part of this recess, which is not preserved, is deeply excavated either directly anterior to, or confluent with, the basioccipital in other non-crocodyliform crocodyliforms and non-crocodylomorph loricatans ( Walker 1990, Gower 2002, Leardi et al. 2020, Ruebenstahl et al. 2022). The preserved ventral body is roughly hourglass-shaped in ventral view, being somewhat constricted in its anteroposterior centre.
The right basipterygoid process is clearly exposed adjacent to its articular surface on the right pterygoid. As preserved, it is composed of two knob-like processes and a large and well excavated concavity lateral to these processes ( Fig. 9C, D View Figure 9 ). In extant crocodylians, as well as crocodyliforms and the hallopodid Almadasuchus figarii , the parabasisphenoid is tightly sutured to the pterygoids and the basipterygoid processes are lost ( Leardi et al. 2020, Kuzmin et al. 2021). However, in several other non-crocodyliform crocodylomorphs the basipterygoid processes form very prominent but hollow protuberances that cover much of the anterior portion of the ventral body in ventral view ( Walker 1990, Leardi et al. 2017, Ruebenstahl et al. 2022). It is possible that much of the ventral surface of the basipterygoid processes of SMNS 12591a are incomplete, and that the large concavity lateral to the two knob-like processes represents the opened internal cavity of the right basipterygoid process. The presence of a cultriform process could not be established, since this region of the parabasisphenoid is covered in ventral view by the pterygoids.
Additional remains of the braincase are visible through the orbits, particularly the right orbit ( Fig. 8A, B View Figure 8 ). They form a transversely thin, plate-like structure that connects to the skull roof dorsally and to the prefrontal anteriorly. The structure is notched along its ventral margin. The remains are poorly preserved and incomplete. Posteriorly, they probably represent part of the laterosphenoid, whereas anteriorly they might be the cristae cranii, which extend far ventrally as a thin lamina from both margins of the olfactory tract of the frontal in Dibothrosuchus elaphros and Junggarsuchus sloani ( Ruebenstahl et al. 2022) .
Mandible
Dentary: Remains of the dentary are preserved in SMNS 12596b, SMNS 12352a, and SMNS 12591a. In SMNS 12596b,
the anterior end of the less dentary is preserved together with a small sliver of the right dentary and they are exposed in ventrolateral view ( Fig. 3B View Figure 3 ). In SMNS 12352a, most of the right dentary is preserved, except for the anteriormost and posteriormost parts. A fragment of the less dentary is also preserved. They are exposed in lateral view and their dorsal margins, as well as their teeth, are mostly obscured by the maxillae ( Fig. 10A–D View Figure 10 ). Both dentaries of SMNS 12591a are virtually completely preserved, only missing the anterior ends ( Figs 8 View Figure 8 , 9C, D View Figure 9 ). However, as for the rest of this skull, the dentaries are strongly dorsoventrally compressed.
The dentary is straight for most of its length, but it widens dorsoventrally and curves slightly dorsally at its anterior end in both SMNS 12596b and SMNS 12352a. This is similar to the condition of Hesperosuchus agilis (AMNH FARB 6758) and Trialestes romeri ( Colbert 1952, Lecuona et al. 2016), whereas the anterior ends of the dentary are not expanded in Terrestrisuchus gracilis , Litargosuchus leptorhynchus , Dromicosuchus grallator , Sphenosuchus acutus , Dibothrosuchus elaphros , and Junggarsuchus sloani ( Crush 1984, Walker 1990, Wu and Chaưerjee 1993, Clark and Sues 2002, Sues et al. 2003, Ruebenstahl et al. 2022). A single, large tooth is preserved on the anterior portion of the less dentary of SMNS 12596b. Two further, more posteriorly positioned, teeth are preserved in the fragmentary right dentary. The dentaries of SMNS 12591a meet at the anterior end of the specimen in ventral view. Since their anteriormost parts are not preserved, the symphysis formed by the dentaries was considerable. The dentary tooth row extended posteriorly until the posterior end of the maxilla but the tooth count cannot be discerned. The posterior articulations of the dentary are unknown because this region is heavily cracked in both mandibular rami in SMNS 12591a.
Splenial: Both splenials are present in SMNS 12591a. The splenial is a thin, plate-like bone that anteriorly contributes to the mandibular symphysis, and posteriorly reaches until the external mandibular fenestra ( Fig. 9C, D View Figure 9 ).
Surangular: Remains of the surangular are exclusively preserved on both mandibular rami of SMNS 12591a ( Figs 8 View Figure 8 , 9C, D View Figure 9 ). Due to extensive crushing its exact outline is unclear but it forms the dorsal margin of the external mandibular fenestra, which is framed ventrally by the angular.The external mandibular fenestra of this specimen was not identified by Ostrom (1981), whereas it was recognized by Huene (1921) and Sereno and Wild (1992). It is badly broken but clearly oval, being anteroposteriorly longer than dorsoventrally tall, as in other crocodylomorphs. The dorsal margin of the surangular is very gently convex.
Angular: Several remains of angulars are preserved in SMNS 12597. Remains of two angulars were identified on SMNS 12597a by Huene (1921), which he considered to probably belong to Procompsognathus triassicus . Crush (1980) also described these elements and referred them to Saltoposuchus connectens . Both elements are incomplete and preserved in close association. The less angular is most complete and exposed in ventral and lateral views, whereas the right angular is partially overlapped by the less element and exposed in medial view ( Fig. 2A View Figure 2 ). A nicely preserved, isolated angular is also preserved in SMNS 12597e ( Fig. 2F View Figure 2 ). This element was originally interpreted as a clavicle by Huene (1921), but it was recognized as an angular by Crush (1980). It represents a largely complete less angular exposed in medial view, and belongs to a different individual than the angulars of SMNS 12597a. Its assignment to Saltoposuchus connectens can therefore only be made tentatively. Both angulars are also present but poorly preserved in SMNS 12591a ( Figs 8 View Figure 8 , 9C, D View Figure 9 ).
The less angular of SMNS 12597a shows that the ventral margin of the angular was gently convex in lateral view and that the angular would have curved around the ventral margin of the mandibular ramus, and thus was visible both in lateral and medial view. The dorsal margin of the element is also visible in lateral view. It was slightly concave, particularly in its central portion, where the angular formed the posteroventral margin of the external mandibular fenestra. Posterior to this, the angular widens somewhat in dorsoventral height, before tapering into a slender posterior process, which would have articulated dorsally with the articular, as in Terrestrisuchus gracilis ( Crush 1984) . The lateral surface of the angular is smooth and slightly transversely convex. The medial surface, as seen the isolated less angular of SMNS 12597e, is markedly transversely concave and characterized by a longitudinal groove approximately in its dorsoventral centre. This groove probably forms the Meckelian canal anteriorly, whereas further posteriorly it formed the articular facet for the pre-articular, as in Terrestrisuchus gracilis (NHMUK PV R 7593a, Crush 1980). The slender posterior process, which forms the articulation facet for the articular, curves posterodorsally and medially, and its medial surface is tilted to face predominantly dorsally.
Pre-articular: On either side of SMNS 12591a, near the posterior end of the mandible, an element is projected posteromedially from the medial side of the mandible. Based on their relative positions, these represent pre-articulars that have been somewhat displaced posteromedially ( Fig. 9C, D View Figure 9 ). They were also identified as such by Huene (1921) and Sereno and Wild (1992). They are incomplete on both sides, but form elongate elements with distinctly transversely convex medial surfaces. Due to the compression of SMNS 12591a, the dorsomedial portions of both mandibular rami are obscured and not externally visible. Therefore, the coronoid could not be confidently identified. Furthermore, because their posterior ends are missing, neither mandibular ramus preserves an articular.
Dentition: The dental distribution and, where possible, tooth count estimates of the premaxilla, maxilla, and dentary are outlined above for those respective bones. All teeth of Saltoposuchus are recurved and labiolingually compressed with readily distinguishable carinae. Serrations are present on both the mesial and distal carinae. They are most obvious on the distal (posterior) carina, where they are present along the entire crown. The entire mesial (anterior) carina of smaller teeth is also serrated, but in larger teeth the proximal half of the crown lacks serrations in the maxillary and dentary teeth in SMNS 12352a ( Fig. 11F View Figure 11 ) and SMNS 12591a, which is also the case in Terrestrisuchus gracilis (NHMUK PV R 7557). Some of the larger teeth, as well as some of the smaller posteriormost teeth, have a very widely rounded labial (lateral) surface. Serrations are clearly present on the distal carinae of the maxillary teeth and the large anterior dentary tooth of SMNS 12596b, but poorly preserved serrations on the mesial carina can only be observed in the second maxillary tooth counted from anterior.
In SMNS 12352a, the largest maxillary tooth appears to be the third or fourth counted from anterior, positioned on the most convex portion of the maxillary alveolar margin in lateral view ( Figs 10A, B View Figure 10 , 11F View Figure 11 ). The maxillary teeth anterior to this are considerably smaller, whereas posteriorly the size of the maxillary teeth reduces gradually. Although less obvious, this condition is also present in SMNS 12591a ( Fig. 8 View Figure 8 ). The presence of enlarged teeth on the anterior portion of the maxilla, as well as the presence of an enlarged dentary tooth near the anterior end of the bone, as seen in the less dentary of SMNS 12596b ( Fig. 3B View Figure 3 ), also occurs in the crocodylomorphs CM 29894, Dibothrosuchus elaphros , and Junggarsuchus sloani ( Wu and Chaưerjee 1993, Clark et al. 2000, Ruebenstahl et al. 2022).
Hyobranchial apparatus: Two slender, rod-like elements are preserved lateroventral to each side of the parabasisphenoid in ventral view in SMNS 12591a ( Fig. 9C, D View Figure 9 ). These slightly curved bones represent the paired ceratobranchial I. They are similar in morphology to those of other non- mesoeucrocodylian crocodylomorphs that preserve this feature of the hyobranchial apparatus in some detail: Terrestrisuchus gracilis and Protosuchus richardsoni (NHMUK PV R 7591, Colbert and Mook 1951).
Postcranium
Cervical vertebrae: Huene (1921) and Crush (1980) both identified two articulated vertebrae preserved in SMNS 12597c as cervical vertebrae. Both are exposed in dorsal view and the anterior and beưer preserved vertebra exhibits the pre- and postzygapophyses, neural arch, and right diapophysis ( Fig. 2E View Figure 2 ). The anterior position of the diapophysis indeed suggests that this element represents a posterior cervical vertebra. However, these elements are not very informative, and the best representatives of the cervical vertebrae are two isolated vertebrae, SMNS 12596d–e ( Fig. 4A–D View Figure 4 ). Based on the relative position of the dia- and parapophysis, both represent approximately mid-cervical vertebrae. Huene (1921) provided a similar interpretation for SMNS 12596d, but considered SMNS 12596e to represent a transitional vertebra between the cervical and dorsal series. Crush (1980) identified both elements as cervical vertebrae. The anterior surface of the centrum is exposed in SMNS 12596e ( Fig. 4C, D View Figure 4 ) and the posterior surface in SMNS 12596d ( Fig. 4A, B View Figure 4 ). Both are concave; the cervical vertebrae were therefore amphicoelous. The ventral margin is strongly concave in lateral view; a clear ventral keel is absent. This corresponds to the condition in the mid- to posterior cervical vertebrae of Terrestrisuchus gracilis (NHMUK PV R 7593b), whereas all cervical vertebrae are keeled in Dromicosuchus grallator ( Sues et al. 2003) . Hypapophyses, as occur in extant crocodylians and Junggarsuchus sloani among non-crocodyliform crocodylomorphs (Hoffsteưer and Gasc 1969, Clark et al. 2004), are also absent. In both elements, the parapophysis is positioned near the anteroventral corner of the lateral side of the centrum, whereas the diapophysis is positioned entirely on the neural arch at approximately the dorsoventral level of the posterodorsal end of the centrum in lateral view.
The neurocentral suture can be observed in SMNS 12596e ( Fig. 4D View Figure 4 ), whereas no suture is visible in SMNS 12596d, which could be the result of poor preservation. Two clear laminae are preserved on the neural arches in both specimens. A prezygodiapophyseal lamina (sensu Wilson 1999) is positioned between the prezygapophysis and diapophysis, and an interzygapophyseal lamina is formed between the pre- and postzygapophyses with a very clear and deeply excavated fossa positioned directly dorsomedial to it ( Fig. 4A–D View Figure 4 ). In addition, a poorly developed lamina extends posteroventrally from the diapophysis, the posterior centrodiapophyseal lamina. The elongate pre- and postzygapophyses of both vertebrae are directed anterodorsally and posterodorsally, respectively, with a strong dorsal inclination. The neural spines are tall and slender. In SMNS 12596e, the neural spine expands very slightly distally in lateral view. In SMNS 12596d, the neural spine appears to taper slightly distally, but this is because its posterior margin is incomplete. The distal ends of the neural spines are flat to slightly convex in lateral view, and not transversely expanded.
Dorsal vertebrae: An articulated series of at least seven dorsal vertebrae is preserved in SMNS 12597a but they are largely embedded in matrix and covered by overlying osteoderms, and only the less sides of their neural arches are exposed. Additionally, two broken but largely complete dorsal vertebrae exposed in right lateral view are preserved in SMNS 12597c ( Fig. 2A, E View Figure 2 ). The right side of a well preserved neural arch of a dorsal vertebrae is exposed in ventral view in SMNS 12596c ( Figs 3D View Figure 3 , 12A View Figure 12 ). Finally, two isolated vertebral centra are preserved in SMNS 55009. Based on the relative elongation of these centra, SMNS 55009a represents a dorsal centrum, whereas SMNS 55009d is a proximal caudal centrum ( Fig. 6A, B View Figure 6 ). However, these elements are largely undiagnostic, and their association with the other remains assigned to SMNS 55009 is unclear, and therefore their assignment to Saltoposuchus connectens should be treated as tentative. Nevertheless, the dorsal centrum has a concave ventral margin in lateral view, and a slightly transversely concave lateral surface, which corresponds to the condition seen in the dorsal centrum of SMNS 12597c and that of Terrestrisuchus gracilis , Hesperosuchus agilis (AMNH FARB 6758), Trialestes romeri , Pseudhesperosuchus jachaleri , and Dibothrosuchus elaphros ( Colbert 1952, Bonaparte 1972, Crush 1984, Wu and Chaưerjee 1993, Lecuona et al. 2016).
The isolated centrum SMNS 55009a is the only element to expose the articular surfaces of the centrum. The centrum is amphicoelous despite being strongly compressed mediolaterally. The most complete dorsal vertebra of SMNS 12597c has a clearly separated dia- and parapophysis ( Fig. 2E View Figure 2 ). The laưer is positioned directly dorsal to the neurocentral suture, just anterior to the anteroposterior mid-level of the vertebra, identifying this element as an anterior dorsal vertebra. The diapophysis is badly broken and no clear laminae can be discerned. SMNS 12596c preserves the morphology of the neural arch of a more posterior dorsal vertebra ( Fig. 12A View Figure 12 ). The diapophysis is extended directly laterally from the neural arch and it is widely rounded distally in ventral view. Its ventral surface is thickened centrally, whereas the anterior and posterior margins are formed by thin laminae. The parapophysis is positioned at roughly the same dorsoventral level as the diapophysis, suggesting this is a mid- to posterior dorsal vertebra. The parapophysis is separated from the diapophysis and its transverse process by a distinct concavity in ventral view.
Dorsal ribs: Some rib fragments are preserved in SMNS 12597a and SMNS 12596c. Both blocks also possess a well preserved example of a dorsal rib, including its proximal end. The well preserved rib of SMNS 12597a is exposed in anterior view, whereas that of SMNS 12596c is exposed in posteroventral view ( Figs 2A View Figure 2 , 3D View Figure 3 , 12A View Figure 12 ). Both ribs are dichocephalous, although the tuberculum of the rib of SMNS 12596c is poorly developed, whereas that of SMNS 12597a is elongate and separated widely from the capitulum. This suggests that the laưer represents an anterior dorsal rib and the former a posterior dorsal rib. It seems likely that almost all dorsal ribs were dichocephalous, with the possible exception of the posteriormost few elements, as in Terrestrisuchus gracilis (NHMUK PV R 7562). The anterior dorsal rib of SMNS 12597a bears a thin lamina that extends from its anterior surface. Proximally, this lamina originates where the capitulum and tuberculum meet, and it tapers out a few mm further distally. It is incomplete. The posterior dorsal rib of SMNS 12596c exhibits a more prominent anterior lamina. It originates at the base of the capitulum, and gradually taper distally towards the end of the preserved rib shass.
Sacral vertebrae and ribs: Remains of two poorly preserved sacral vertebrae are associated with the pelvic girdle and dorsal vertebral series in SMNS 12597a ( Figs 2A View Figure 2 , 12C View Figure 12 ). Huene (1921) suggested three sacral vertebrae were present here, but Crush (1980) correctly identified the presence of only two sacrals, as in all other crocodylomorphs, except for rare cases in which a dorsal or caudal vertebra is incorporated in the sacrum ( Scheyer et al. 2019). A single, disarticulated sacral vertebra is preserved in SMNS 12596c and in SMNS 55009b ( Figs 3C, D View Figure 3 , 4E, F View Figure 4 , 6C, D View Figure 6 ). The sacral remains of SMNS 12597a are represented by the right sacral ribs exposed in dorsal view and are preserved in articulation with the right ilium ( Fig. 12C View Figure 12 ). Crush (1980) also identified a part of the less rib of the second sacral vertebra, but this element is not recognized here. The ribs of both vertebrae are strongly developed and anteroposteriorly broaden distally. The second sacral rib is broader than the first. The first sacral rib has concave anterior and posterior margins in dorsal view, with the former being most concave. This rib is most expanded anterodistally. Its dorsal surface is mostly transversely concave, which might be the result of compression, apart from a raised, flaưened platform on its anterodistal portion. The second sacral rib is composed of an elevated anterior half and a posterior half that is deflected posteroventrally. Both portions are separated by a proximodistally directed, thin ridge, and the dorsal surfaces of both portions are gently transversely concave. The anterior margin is concave, whereas the posterior margin is incomplete.
The sacral vertebra of SMNS 12596c is a second sacral vertebra exposed in right lateral view ( Fig. 4E, F View Figure 4 ). The neural arch, including the neural spine and right pre- and postzygapophysis are preserved, as well as the right rib. The less rib of the same element can be observed on the other side of the slab ( Fig. 3C View Figure 3 ). The postzygapophysis is placed considerably further dorsally than the prezygapophysis. The neural spine is low and its lateral surface is distinctly transversely convex. An incomplete interdigitating suture is present between the transverse process and rib; it is placed far proximally, near the base of the neural spine. The right rib broadens distally and both the anterior and posterior margins are concave. The dorsal surface of the rib is transverse concave. Distally, the rib is strongly expanded posteriorly to form a dorsoventrally thin posterior flange, similar to that seen in Terrestrisuchus gracilis , Hallopus victor , and Dromicosuchus grallator ( Walker 1970, Crush 1984, Sues et al. 2003). However, a distinct groove on the lateral surface of the rib associated with the flange, as present in Terrestrisuchus gracilis , is absent. The lateral surface of the rib is rugose and, except for the thin posterior flange, it is dorsoventrally thickened with a transversely gently concave lateral surface. The dorsal side of the posterior flange is convex in lateral view, and its ventral side concave.
The sacral vertebra of SMNS 55009b is strongly deformed so that the less side of the specimen, including the less rib, is displaced ventrally and the right side dorsally ( Fig. 6C, D View Figure 6 ). The exposed anterior articular surface is clearly concave. The neural spine has a gradual transverse expansion distally but its lateral margin is flat to slightly concave, although it is unclear whether this is the result of compression. The postzygapophysis is again positioned considerably further dorsally than the prezygapophysis and an interdigitating suture is present between the rib and transverse process near the base of the neural spine. The rib is badly broken laterally and ventrally but clearly broadens distally in dorsal view. Its dorsal surface is mostly flat. The lateral side of the rib is incomplete and largely uninformative, but a thin posterior flange appears to be lacking; this element is therefore identified as a first sacral vertebra (sensu Crush 1980 but contra Huene 1921, who identified it as a second sacral vertebra). Due to its isolated nature and lack of diagnostic features, the assignment of this element to Saltoposuchus connectens is only tentative.
Caudal vertebrae and ribs: An articulated series of both the proximal and distal caudal vertebrae is preserved in SMNS 12597. The proximal series is preserved in SMNS 12597a and the distal series in SMNS 12597d ( Fig. 2B, D View Figure 2 ). The slabs do not connect directly and, based on the presence of a well developed transverse process on the posteriormost preserved caudal vertebra of SMNS 12597a, and the complete absence of a transverse process on the anteriormost caudal vertebra on SMNS 12597d, a considerable part of the tail is missing, as was also suggested by Huene (1921). A partially preserved, isolated caudal vertebra is present in SMNS 12596f. It is exposed in less lateral view, and based on the presence of a poorly developed transverse process, this represents approximately a vertebra from the middle of the caudal series ( Fig. 4G, H View Figure 4 ). Two further, very poorly preserved, proximal caudal vertebrae are preserved in SMNS 12596h in association with a less ischium. They are exposed in right lateral view and the complete right side of the neural arch dorsal to the transverse process is visible in the proximal element, whereas the right prezygapophysis and a small part of the centrum is visible for the distal vertebra ( Fig. 5C View Figure 5 ). SMNS 55009d represents an isolated centrum of a proximal caudal vertebra and SMNS 55009c represents a complete and isolated, proximal caudal vertebra including ribs exposed in posterior view ( Fig. 6B, E, F View Figure 6 ).
Nine articulated, proximal caudal vertebrae are preserved on SMNS 12597a, whereas 15 distal caudal vertebrae are preserved in SMNS 12597d ( Fig. 2B, D View Figure 2 ). The end of the tail is not preserved, but the diminutive size of the posteriormost caudal vertebrae indicates that they are positioned very close to the terminus. Based on the separation between the sacrum and proximalmost exposed caudal vertebra in SMNS 12597a, as well as the presence of a chevron anterior to the anteriormost exposed caudal vertebra, at least one but possibly a few caudal vertebrae are still imbedded in matrix, as was also suggested by Huene (1921). Huene (1921: fig. 31) estimated that the gap between both blocks comprised at least 15 vertebrae and reconstructed Saltoposuchus connectens with 53 caudal vertebrae, which is very likely an exaggeration. Protosuchus richardsoni is the only early crocodylomorph for which the caudal vertebral count is known, possessing around 35 caudal vertebrae ( Colbert and Mook 1951), whilst in modern crocodylians the caudal vertebral count is 30 to 40 (Hoffsteưer and Gasc 1969). It seems likely that the count of Saltoposuchus connectens also falls within this range.
The neurocentral suture is unfused in both the proximal and distal caudal vertebrae of SMNS 12597, suggesting this specimen is likely immature. The suture between the transverse process and caudal rib is visible in some but not all proximal caudal vertebrae. The ventral margin of all caudal centra is very gently concave. The posterior articular surface of SMNS 55009c is slightly concave, whereas the deeply excavated articular faces of SMNS 55009d are likely exaggerated due to compression ( Fig. 6B, E, F View Figure 6 ). The relative elongation of the caudal centra increases considerably posteriorly. The transverse processes and ribs are oriented perpendicular to the axis of the vertebral column. They do not extend particularly far laterally and have a rounded to rectangular distal end. Chevrons articulate between the ventral edges of the caudal vertebrae ( Fig. 2B, D View Figure 2 ). They are directed strongly posteriorly and only slightly ventrally, and have a posterodorsal curvature in lateral view. They occur along the entire tail, including the distalmost vertebrae, but reduce in length gradually posteriorly along the series.
The neural spine of the isolated mid-caudal vertebra SMNS 12596f is slightly directed anterodorsally in lateral view ( Fig. 4G, H View Figure 4 ). Anterior to the neural spine, an additional, sharply pointed, anterodorsally directed process is present, which is separated from the spine by a gap that is widely rounded in lateral view. This is the accessory anterior spine (sensu Clark 1986: ch. 14), which is also present in Terrestrisuchus gracilis (NHMUK PV R 7561) but absent in Hallopus victor and Sphenosuchus acutus among non-crocodyliform crocodylomorphs ( Walker 1970, 1990, Crush 1984). Neural spines are also present in the two caudal vertebrae of SMNS 12596h, but these are poorly preserved and incomplete anteriorly, and only the base of the accessory anterior spine is preserved ( Fig. 5C View Figure 5 ). In the distal caudal vertebrae, transverse processes are absent and the neural spines are very strongly reduced, but their morphology cannot be observed because they are covered dorsally by osteoderms in SMNS 12597e ( Fig. 2C View Figure 2 ).
Osteoderms: Many osteoderms are preserved in SMNS 12597a, d, e and SMNS 12596c ( Figs 2A, B, D, E View Figure 2 , 3C, D View Figure 3 , 4E, F View Figure 4 , 12A–C View Figure 12 ). In SMNS 12597, they form a paired row across both the dorsal and caudal vertebral column. Only the osteoderms on the less side of the dorsal column are well preserved, but slivers of the osteoderms of the right row can also be discerned ( Fig. 2A View Figure 2 ). On the tail the osteoderms reach until the posteriormost preserved vertebra ( Fig. 2D View Figure 2 ). It is unclear whether the osteoderms are also paired here. Huene (1921) identified a tiny, lancet-shaped osteoderm associated with the two cervical vertebrae of SMNS 12597c. In contrast to Crush (1980), I have recovered this element, but I do not agree with its identification. It most likely represents part of the transverse process of a vertebra. Thus, no cervical osteoderms are known for Saltoposuchus connectens , but they were almost certainly present, as in other crocodylomorphs. Appendicular and ventral osteoderms are absent, as in all other known non-crocodyliform crocodylomorphs. The osteoderms differ in size along the vertebral column but maintain a similar morphology. The osteoderms along the dorsal vertebral series of SMNS 12597a gradually increase in width and level of ornamentation posteriorly, whereas on the tail they decrease in size posteriorly ( Fig. 2A, D View Figure 2 ).
The dorsal surface of the osteoderms is heavily sculptured by pits and ridges and, additionally, possesses a very low and narrow longitudinal ridge that is placed slightly medial to the transverse midpoint of the osteoderm ( Figs 4E, F View Figure 4 , 12B View Figure 12 ). The osteoderms are subrectangular to oval in outline in dorsal view, being considerably longer anteroposteriorly than mediolaterally wide. The medial margin, which articulated with its antimere, is straight, whereas the lateral margin is convex in dorsal view. The osteoderms of Saltoposuchus connectens are similar to those seen in Dromicosuchus grallator ( Sues et al. 2003) , relatively much larger and much more sculpted than the osteoderms of Terrestrisuchus gracilis ( Crush 1984) , and less rectangular but rather more oval-shaped than the osteoderms of Hesperosuchus agilis (AMNH FARB 6758) and CM 29894 ( Colbert 1952, Clark et al. 2000). They possess a short, anteriorly directed peg on their anterior margin as seen in disarticulated osteoderms of SMNS 12596c. The ventral surface, exposed in an osteoderm directly adjacent to the less transverse process of the sacral vertebra in SMNS 12596c, is smooth and gently transversely concave and its posterior margin possesses a distinct notch that would have received the anterior peg of the subsequent osteoderm.
Scapula: The distal portion of a right scapular blade is preserved in SMNS 12597a ( Figs 2A View Figure 2 , 12D View Figure 12 ). Its exposed lateral surface is smooth and flaưened to slightly transversely convex. The distal end is expanded both anteriorly and posteriorly. The posterior expansion is more gradual and positioned further dorsally than the anterior expansion and, consequently, the anterior margin is more strongly concave than the posterior one. Both expansions terminate in an acute tip, which is sharpest in the posterior expansion. The outer surface of the dorsal margin is incomplete, which according to Crush (1980) represents an area of partial ossification. Regardless, its outline is clearly preserved. It is distinctly convex in lateral view and from its apex it gradually curves posteroventrally towards the tip of the posterior expansion and anteroventrally under a stronger inclination towards the tip of the anterior expansion. The outline of the distal end of the scapula is very similar to that of Terrestrisuchus gracilis and Hesperosuchus agilis (AMNH FARB 6758, Colbert 1952, Crush 1984), contrasting with the anteroposteriorly wider scapular blades in Sphenosuchus acutus , Trialestes romeri , Dibothrosuchus elaphros , and Junggarsuchus sloani ( Walker 1990, Wu and Chaưerjee 1993, Clark et al. 2004, Lecuona et al. 2016). A small fragment preserved dorsally to the dorsal end of the scapula was tentatively identified as part of the scapulacoracoid by Huene (1921). This fragment is considered unidentifiable here.
Ulna: A long bone present among SMNS 12596 was identified by Crush (1980) and Hughes (1963) as an ulna, asser originally being interpreted as a metatarsal by Huene (1921). Based on its length (Supporting Information, Table S1), it is too long to represent a metatarsal. One end of the element is asymmetrical, with one of its sides reaching further away from the shass, which could correspond to the olecranon process on the proximal end of an ulna ( Fig. 5D View Figure 5 ). A small process projecting from the side of the head might correspond to the anteromedial tuber, which would suggest that the element represents a less ulna. However, other aspects of the element differ from known long bones of early crocodylomorphs; the shass is curved, flaưened, and widened towards its distal end, which could potentially be the result of a pathology or diagenetic compression, and the inferred distal end of the bone is very thin. The presence of one asymmetrical end and another flaưened and widened end also does not correspond with the morphology of a metatarsal. Its identification as a less ulna of Saltoposuchus connectens is tentatively maintained here, although, due to the disassociation of the material, it cannot be excluded that the element belongs to a different taxon.
Carpus and manus: An articulated less manus is preserved and exposed in dorsal and medial views in SMNS 12352b ( Fig. 8E–H View Figure 8 ). Directly proximal to metacarpals I and II, the distal half of an elongate element is preserved. This element was previously identified as the less radius ( Huene 1921, Ostrom 1981), but Sereno and Wild (1992) later identified this element as a partial, elongate less radiale. They also identified a wide but thin piece of bone between the radiale and metacarpal II as a distal carpal, and a tiny bone sliver lateral to the distal end of the radiale as the distalmost end of the less ulnare. This configuration of a single distal carpal and an elongate radiale and ulnare proximal to this distal carpal is corroborated here, and forms a diagnostic configuration for early crocodylomorphs ( Crush 1984, Wu and Chaưerjee 1993, Nesbiư 2011: ch. 242, Lecuona et al. 2016). In contrast, it does not correspond to the carpal configuration of early theropods, in which the radiale and ulnare are very wide and proximodistally very short elements, and in which three distal carpals are generally present (e.g. Colbert 1989, Nesbiư 2011). The manus and associated elements therefore clearly belong to a non-crocodyliform crocodylomorph (sensu Sereno and Wild 1992), and they can reasonably be assigned to Saltoposuchus connectens based on their association with the skull of SMNS 12352a. The shass of the radiale has a flaưened anterior face and is slightly wider mediolaterally than anteroposteriorly. The distal end of the radiale is slightly mediolaterally expanded compared to the shass. The ulnare preserves no discernible features, whereas the distal carpal is also poorly preserved but roughly ovoid in outline. It is only slightly wider than the proximal head of metacarpal II.
All metacarpals are virtually completely preserved.Metacarpal I is considerably shorter, but not narrower, than the other metacarpals ( Fig. 10E–H View Figure 10 ), as was also observed previously ( Huene 1921, Ostrom 1981, Sereno and Wild 1992). The other metacarpals are all elongate, with metacarpal V being the shortest, followed by metacarpals IV and then II, and with metacarpal III being the longest (Supporting Information, Table S1). The metacarpals are subequal in width, except for metacarpal V, which is more slender. The metacarpals are all tightly bunched and metacarpals II–V are all oriented in the same lateromedial plane, whereas metacarpal I is positioned ventromedial to metacarpal II ( Fig. 10G, H View Figure 10 ). The tightly bunched carpus is indicative of a digitigrade posture, as was previously also pointed out by Sereno and Wild (1992) for Saltoposuchus connectens . A digitigrade posture of the front limb has also been described for Terrestrisuchus gracilis, CM 29894, Junggarsuchus sloani , and Trialestes romeri , albeit tentatively for this final taxon (NHMUK PV R 7562, NHMUK PV R 7557b, Clark et al. 2000, 2004, Lecuona et al. 2016). The metacarpals are slightly expanded at their proximal and distal ends, and the dorsal surface of the distal ends possess concave extensor pits ( Fig. 10E, F View Figure 10 ). Two phalanges are preserved in digit I, of which the distalmost represents a very small ungual ( Fig. 10G, H View Figure 10 ). No further unguals are preserved and the phalangeal count of the other digits is incomplete. Digits II, IV, and V each preserve two phalanges and digit III preserves three ( Fig. 10E, F View Figure 10 ). Extensor pits are also present on the distal ends of the phalanges.
Ilium: A single, right ilium is preserved in articulation with the sacral ribs and femur in SMNS 12597a ( Figs 2A View Figure 2 , 12C View Figure 12 ). It is poorly preserved and exposed in dorsal view. The pre-acetabular process is largely missing but, although its dorsal margin is missing, the post-acetabular process is preserved across its entire length. The dorsal margin of the ilium has a sinusoidal outline in dorsal view, with the post-acetabular process curving first medially and subsequently laterally from anterior to posterior ( Fig. 12C View Figure 12 ). This is probably due to diagenetic deformation. The lateral surface of the ilium slants ventrolaterally to form the supracetabular crest, which extends far laterally beyond the rest of the ilium. Its lateral margin is incomplete. The acetabulum itself is covered by the crest and completely embedded in matrix. The medial side of the ilium is largely covered in matrix, but it is apparent that the first sacral rib aưaches on the anterior portion of the main body of the ilium, with its anterior end connecting to the now missing pre-acetabular process. The second sacral rib connects on its anterior end to the posterior part of the main body of the ilium and on its posterior part to the proximal base of the post-acetabular process.
Ischium: A large right ischium is exposed in medial or dorsal view in SMNS 12596h ( Fig. 5C View Figure 5 ). It is well preserved but its ventral, anterior, and probably posterior margins are incomplete and therefore the proximodistal length of the ischial shass cannot be determined. The ischial shass curves posteriorly so that its distal end is directed considerably more posteriorly than ventrally. The ischial shass gradually reduces in height distally. The dorsal margin is distinctly concave in medial view, thickened and curved proximally, but thin distally. From the base of the dorsal end of the ischial shass a crest is projected that curves anterodorsally and medially. At its dorsal end, this crest forms the posterior end of a large, concave surface that faces mediodorsally and somewhat anteriorly and that is triangular in outline. This forms the articular surface with the ilium. The medial surface directly ventral to the iliac peduncle is concave. A small, sloped surface is visible directly lateral to the peduncle, which is directed dorsolaterally and slightly anteriorly. This probably represents the incompletely exposed antitrochanter of the ischium (contra Sereno and Wild 1992), which is also present in Terrestrisuchus gracilis but absent in Trialestes romeri (NHMUK PV R 37728, Lecuona et al. 2016). A clear concavity anterior to the iliac peduncle, as is found in Terrestrisuchus gracilis (NHMUK PV R 37728), is absent, which could suggest that the acetabular fenestra was comparatively smaller in Saltoposuchus connectens . The incomplete anterior margin is roughly straight and vertically oriented. This formed the articular surface with the pubis. The incomplete ventral margin formed the articular surface for the less ischium.
Pubis: The pubis is preserved in SMNS 12597a and SMNS 12596g. The right pubis of SMNS 12597a is disarticulated from, but still associated with, the right ilium and sacral vertebrae. Most of the shass is exposed in lateral and anterior views ( Fig. 2A View Figure 2 ). SMNS 12596g preserves a virtually complete, disarticulated right pubis that is exposed in anterior and medial views ( Fig. 5A, B View Figure 5 ). Its proximal portion is composed of a plate-like surface that is directed ventromedially to dorsolaterally, with the dorsal end being curved so that it is oriented directly dorsally. The plate is characterized by a very large and oval-shaped obturator foramen, which has a long axis that is directed anteroventrally to posterodorsally. The posterior margin of the pubic plate is composed of a dorsal portion, which forms approximately two-thirds of the posterior margin, and a ventral portion. The dorsal portion of the posterior margin is concave in medial view, and forms the articular surface with the ischium, whereas the ventral portion is straight and did not articulate with another element, thus forming an open notch. In contrast, the straight ventromedial margin of the plate is aligned with the medial margin of the pubic shass, and both would have articulated with the opposing pubis. The dorsal margin of the pubic plate in medial view is incomplete but would have articulated with the ilium. Near its proximal end, the pubis possesses a clear laterally and slightly posteriorly directed process, which has a triangular outline in anterolateral view. This is the pubic tubercle, which is also present in Terrestrisuchus gracilis and Trialestes romeri (NHMUK PV R 7562, Lecuona et al. 2016), and which forms an aưachment site for M. obliquus abdominus, M. ambiens, and ligaments ( Hutchinson 2001b). The portion posterodorsal to the pubic tubercle represents the iliac peduncle, which is embedded in matrix and largely obscured by an overlying osteoderm.
The pubis possesses an elongate, flaưened pubic shass that has wide anterodorsal and posteroventral surfaces and a thin and incomplete medial margin and a somewhat thickened lateral margin. The torsion between the proximal body of the pubis and the pubic shass is greater than in Terrestrisuchus gracilis (NHMUK PV R 7561b, Crush 1984), such that the obturator foramen cannot be observed in anterior view. There is also some torsion in the pubic shass itself, so that its anterior surface curves very slightly laterally towards the distal end. The pubic shass of SMNS 12596g is incomplete distally. The lateral margin of the shass gently curves mediodistally so that it very gradually reduces in transverse width distally. The shass is also gently curved anteroventrally, so that its anterior margin is convex in lateral or medial view.
Femur: Both femora are virtually completely preserved but damaged in SMNS 12597a ( Fig. 2A View Figure 2 ). The right femur is preserved in association with the right ilium and sacral vertebrae proximally, and the right tibia and fibula distally, although the laưer elements have been displaced distally. The proximal end of the less femur has been displaced from the vertebral column and is positioned adjacent to the angulars, but distally it is found in complete articulation with the less tibia and fibula. Both femora of SMNS 12596 (c, j) are also preserved ( Figs 3C View Figure 3 , 5E–I View Figure 5 ), and they are in beưer condition than those in SMNS 12597a. The less femur of SMNS 12596c is preserved in association with disarticulated vertebrae, ribs, and osteoderms. The right femur is completely freed from matrix, and based on its length and congruent morphology belongs to the same individual (Supporting Information, Table S1). All femora have been considerably transversely compressed.
As noted by previous studies ( Crush 1980, Irmis et al. 2013), the femur of Saltoposuchus connectens is considerably more robust (i.e. with a comparatively broad circumference relative to total length) than that of Terrestrisuchus gracilis ( Irmis et al. 2013: 284) . The femur is longer than the tibia and fibula ( Table 2; Sereno and Wild 1992) as in Dromicosuchus grallator and Pseudhesperosuchus jachaleri ( Sues et al. 2003, Leardi et al. 2017: supplementary information therein), whereas in Terrestrisuchus gracilis , Hallopus victor , and Macelognathus vagans the tibia and fibula are longer than the femur (NHMUK PV R 7551, Walker 1970, Göhlich et al. 2005). The femoral shass is distinctly curved in anterolateral or posteromedial view, so that the anteromedial margin of the shass is clearly convex and the posterolateral margin clearly concave ( Fig. 5E, F View Figure 5 ).
The proximal femoral head is strongly inturned and directed anteromedially to articulate with the acetabulum. The proximal surface of the proximal head is convex and gradually narrows anteromedially to posterolaterally ( Fig. 5G View Figure 5 ). Of the three tubera typically present on the proximal head of the femur in archosaurs ( Nesbiư 2011: chs 300–302), only a poorly developed anteromedial tuber can be observed on the right femur SMNS 12596j due to compression. SMNS 12597a exposes a clearly defined posteromedial tuber on its less femur and a similarly defined anterolateral tuber on its right femur. The posterolateral margin bears a longitudinal, rugose thickening directly distal to the proximal head. It is positioned on the lateral side of the margin and represents the lesser or dorsolateral trochanter ( Fig. 5F, I View Figure 5 ), which occurs widely among non-crocodyliform crocodylomorphs and represents an aưachment site for M. puboischiofemoralis internus 2 (NHMUK PV R 7562, Hutchinson 2001a, Sues et al. 2003, Göhlich et al. 2005, Pol et al. 2013). The fourth trochanter is low and mount-like and the surface of the shass at its anteromedial base is quite concave ( Fig. 5E, I View Figure 5 ).
The compressed distal end of the right femur of SMNS 12596j bears distinct fibular and tibial condyles, which are positioned on the lateral and medial side of the distal end, respectively ( Fig. 5H View Figure 5 ). They are separated by a distinct groove on the distal surface that curves proximally onto the anterior surface of the shass where it continues proximally for a considerable length. The two condyles are separated by a strong concavity on the posterior margin of the distal end. In distal view, the lateral condyle is considerably narrower and reaches further posteriorly than the medial condyle. Although this is strongly exaggerated by compression, this would have probably also been the case in life to a lesser extent, as can be deduced from the well preserved femur of Terrestrisuchus gracilis (NHMUK PV R 37685). A large crista tibiofibularis is projected from the lateral side of the lateral condyle. It is separated from the posterior end of the lateral condyle by a distinct notch.
Tibia and fibula: Both pairs of tibiae and fibulae are preserved in articulation in SMNS 12597a. The right pair lacks the distal ends of both elements, whereas the less pair is complete, although the central portion of the fibular shass is completely covered in matrix. The less pair is preserved in articulation with the less femur proximally and less tarsus distally ( Fig. 2A, B View Figure 2 ). A less tibia and fibula are fully freed from matrix and preserved in partial articulation in SMNS 12596k ( Fig. 5J, K View Figure 5 ). Finally, SMNS 55009e is a complete, well preserved right tibia that is fully freed from matrix ( Fig. 6A–F View Figure 6 ). The tibia and fibula are subequal in length, but the tibia is considerably more robust (Supporting Information, Table S1). The proximal ends of both elements are curved relative to the shass, so that the posterior margin directly distal to the proximal end is concave, and the opposite anterior end convex. The fibula of SMNS 12596k is apparently shissed relative to the tibia, so that its concave posterior margin faces the equivalent margin on the tibia ( Fig. 5J, K View Figure 5 ). Distally, both elements also gradually curve anteriorly and as such have a somewhat sigmoidal outline. The curvature of the tibia SMNS 55009e is much reduced due to its strong anteroposterior compression ( Fig. 7A–D View Figure 7 ).
The proximal surface of the tibia is slightly concave. The anterior margin of the proximal head is widely rounded and thus convex in proximal view, whereas the posterior side possesses two projections, which represent the posterior expansion on the posteromedial end and the lateral condyle on the posterolateral end respectively ( Figs 7E View Figure 7 , 12E View Figure 12 ). The posterior expansion extends quite far beyond the shass in all tibiae of Saltoposuchus connectens , and a similar projection is also present in Terrestrisuchus gracilis and Dromicosuchus grallator (NHMUK PV R 7566, Sues et al. 2003). The markedly exaggerated expansion in SMNS 55009e is the result of compression. The posterior expansion and lateral condyle are separated by a deep concavity in proximal view, and this concavity continues distally along the posterior surface of the shass for some distance as a trough ( Fig. 7A View Figure 7 ). The lateral side of the proximal head also possesses a small but distinct notch in proximal view ( Crush 1980; Figs 7E View Figure 7 , 12E View Figure 12 ) and such a notch is absent in Terrestrisuchus gracilis (NHMUK PV R 7566, Crush 1984). The distal end of the tibia is composed of two distal projections, one on the anterolateral side and the other on the posteromedial side ( Fig. 7A, B View Figure 7 ). In other non-crocodyliform crocodylomorphs the posteromedial side projects further distally than the anterolateral side (NHMUK PV R 7566, Colbert 1952, Sues et al. 2003, Göhlich et al. 2005). In both SMNS 55009e and SMNS 12596k, this condition is reversed, with the anterolateral side extending further distally ( Figs 5J View Figure 5 , 7A, B View Figure 7 ). This might represent an artefact of compression, since this extension is strongly exaggerated in SMNS 55009e, and the tibia of SMNS 12596k is also compressed.
The proximal and distal ends of the fibula are very slightly expanded relative to the shass and due to compression they form a thin ridge rather than a clear surface in SMNS 12596k ( Fig. 5J, K View Figure 5 ). The anterolateral side of the fibula possesses a thin crest on the proximal half of the shass ( Fig. 5K View Figure 5 ). This crest is demarcated by a marked groove on the lateral surface of the shass posterior to it, which might also represent a compression artefact. This anterolateral crest (=anterior trochanter sensu Sereno 1991) was previously described by Huene (1921) as an aưachment site for M. iliofibularis.
Tarsus: Of the tarsus, three calcanea and a distal tarsal 4 are preserved. The less calcaneum and distal tarsal 4 are preserved in SMNS 12597a in association with the less tibia and fibula and the less pes, whereas the right calcaneum SMNS 12597f is completely freed from matrix ( Figs 2B, G, H View Figure 2 , 12F, G View Figure 12 ). Finally, SMNS 55009g is another right calcaneum completely freed from matrix ( Fig. 7J–L View Figure 7 ). The less calcaneum of SMNS 12597a exposes its flaưened lateral surface, including a posteriorly directed calcaneal tuber that curves posteroproximally and which has a distinctly concave proximal surface ( Fig. 12G View Figure 12 ). A posteriorly directed calcaneal tuber is indicative of an erect posture (sensu Parrish 1987) and also occurs in the non-crocodyliform crocodylomorphs Hallopus victor , Trialestes romeri , Terrestrisuchus gracilis, YPM 41198, and Macelognathus vagans ( Parrish 1987, Göhlich et al. 2005, Nesbiư 2011, Lecuona et al. 2016). It deviates from the more posterolaterally oriented calcaneal tubera of extant crocodylians ( Parrish 1987). The anterior margin of the calcaneum is curved anteriorly and proximally, whereas its anterodistal portion is flattened and in articulation with the distal tarsal 4 directly distal to it. A process is projected proximal to the main body of the calcaneum. This probably represents the heavily crushed lip from the medial side of the calcaneum, which has been shissed proximally, whereas it was projected medially in life ( Fig. 12G View Figure 12 ). Anteriorly, the lip is deeply excavated, and here it formed the posterior surface of the deep facet that received the laterally projected peg of the astragalus ( Sereno 1991).
The element distal to this calcaneum was interpreted as the astragalus by Huene (1921), but it was later identified as distal tarsal 4 by Crush (1980). It is around two-thirds the size of the calcaneum ( Fig. 12G View Figure 12 ). It is widest at its posterodistal end relative to the calcaneum, and narrower at its anteroproximal end. It also possesses a small, laterally directed process near its proximal end. Its morphology is somewhat similar to that of distal tarsal 4 of pseudosuchian-line archosaurs, including YPM 41198, which also possesses a narrow and wide end, respectively ( Nesbiư 2011: ch. 349). Furthermore, the element lacks the complex structure seen in the astragalus of crocodylomorphs that includes a prominent peg for the articulation with the calcaneum ( Sereno 1991). The interpretation of this element as a distal tarsal 4 is therefore corroborated.
The isolated right calcaneum of SMNS 12597f possesses a complete calcaneal tuber that roughly corresponds in morphology to that of the less calcaneum ( Fig. 2G, H View Figure 2 ). Its main body is badly broken, with its proximal half missing. Nevertheless, its medial surface was clearly deeply excavated, forming the facet for the astragalar peg. The right calcaneum SMNS 12597f was already isolated from the rest of the material of SMNS 12597 in 1921 ( Huene 1921). This element is considerably larger than the associated less calcaneum of SMNS 12597a, to such an extent (Supporting Information, Table S1) that it is certain that both elements derive from separate individuals. Furthermore, the right calcaneum SMNS 12597f is also stouter and less anteroposteriorly elongate compared to the calcanea of SMNS 12597a and SMNS 55009g ( Figs 2G, H View Figure 2 , 7I–L View Figure 7 , 12G View Figure 12 ). This could be related to allometric variation or an anteroposterior compression of the element. Based on the prominent calcaneal tuber and deeply excavated astragalar facet that is framed posteriorly by a distinct lip, this element clearly belongs to a crocodylomorph and its assignment to Saltoposuchus connectens is tentatively maintained. The isolated right calcaneum SMNS 55009g is mostly complete, although the distal surface of the calcaneal tuber is missing, and it is poorly preserved, being heavily distorted ( Fig. 7I–L View Figure 7 ). It possesses the same morphology as that seen in the calcanea of SMNS 12597, as well as those of other crocodylomorphs.
Pes: Remains of the less pes are preserved in SMNS 12597a in articulation with the rest of the less hindlimb. They consist of very fragmentary remains of at least two metatarsals and the phalanges of the corresponding digits ( Figs 2B View Figure 2 , 12F–H View Figure 12 ). Only the proximal ends of the metatarsals are informative. The outer element is more gracile than the element associated with distal tarsal 4, and its proximal head is smaller and bulbous ( Fig. 12F, G View Figure 12 ). This represents metatarsal V, as was also suggested by Huene (1921). The proximal head of the second metatarsal is incomplete but markedly broader than that of metatarsal V.
Three incomplete metatarsals are preserved in SMNS 55009f ( Fig. 7G, H View Figure 7 ). Their shasss are straight, elongate, and tightly bundled, similar to the condition in Terrestrisuchus gracilis , Hallopus victor , and Macelognathus vagans (NHMUK PV R 7551b, Walker 1970, Crush 1984, Göhlich et al. 2005), whereas they are slightly more spread in Trialestes romeri ( Lecuona et al. 2016) . The metatarsals differ from other non-crocodyliform crocodylomorphs in possessing a distinct longitudinal groove along the shass. This was aưributed to compression by Crush (1980) but it cannot be excluded that such a groove, possibly to a lesser extent, was also present in life.
The phalanges of SMNS 12597a have been partially disarticulated, preventing their referral to specific digits and determining the phalangeal count ( Fig. 12F, H View Figure 12 ). However, based on the presence of two more robust, proximally positioned phalanges and two unguals, they likely represent remains of two digits. The non-terminal phalanges possess expanded proximal and distal ends relative to the shass. The proximal ends are flat to slightly concave, whereas the distal ends are marked by distinct extensor pits. The unguals are small in size and curved in lateral view ( Fig. 12H View Figure 12 ).
Histological section of the right femur of SMNS 12596
The strong anterolateral to posteromedial compression of the right femur SMNS 12596j is also reflected in the histological section of its mid-shass. The medullary cavity has been diagenetically reduced to an anteromedially to posterolaterally oriented slit-like opening ( Fig. 13A View Figure 13 ). Along the preserved long axis, the medullary cavity comprises approximately half of the diameter of the shass. Previously studied long bone sections of non-crocodyliform crocodylomorphs comprise an isolated, likely immature humerus assigned to Terrestrisuchus sp. (Riqclès et al. 2003) and a femur and long bone fragments of Hesperosuchus agilis (AMNH FARB 6767, material associated with holotype specimen AMNH FARB 6758; Riqclès et al. 2008, Buffrénil et al. 2021). In the former, the medullary cavity is clearly considerably larger compared to the preserved long axis in SMNS 12596j, whereas the medullary cavity of the laưer is approximately similar in width. However, it is largely filled by extensive trabeculae in Hesperosuchus agilis , which are absent in both Terrestrisuchus sp. and Saltoposuchus connectens . In SMNS 12596j, the inner margin of the periosteal tissue has been resorbed and subsequently an endosteal layer of lamellar bone with flaưened and elongated osteocyte lacunae has been deposited that is strongly birefringent in polarized light ( Fig. 13B, C View Figure 13 ). This layer is considerably thicker than the very thin endosteal layer identified in the humerus of Terrestrisuchus sp. (Riqclès et al. 2003).
The periosteal matrix of SMNS 12596j is composed of primary fibrolamellar or woven-parallel bone with numerous, relatively round osteocyte lacunae. The cortex possesses a high density of simple vascular canals that are roughly equally distributed throughout the matrix ( Fig. 13B, C View Figure 13 ). No secondary osteons are present, in contrast to the femoral section of Hesperosuchus agilis (Buffrénil et al. 2021) . The vascular canals are predominantly longitudinally oriented, with some forming anastomoses ( Fig. 13A View Figure 13 ). As such, the composition of the periosteal matrix is broadly similar to that seen in the humerus of Terrestrisuchus sp. (Riqclès et al. 2003). In contrast, in Hesperosuchus agilis the periosteal matrix is formed by parallel-fibred tissue that is very distinctly lamellar for most of its width, being least organized near the medullary cavity (Riqclès et al. 2008, Buffrénil et al. 2021). In the humerus of Terrestrisuchus sp. the periosteal matrix is most organized (i.e. lamellar) near the periphery, as is also indicated by the presence of the two lines of arrested growth (LAGs) (Riqclès et al. 2003). Sharpey’s fibres, as present in Hesperosuchus agilis (Buffrénil et al. 2021) , are absent in SMNS 12596j. Two regions of increased organisation are present in the periosteal matrix of SMNS 12596j, most clearly visible in polarized light. Each area is composed of three closely packed lines ( Fig. 13D, E View Figure 13 ), which might reflect temporarily arrested growth. The first region of arrested growth is formed at approximately the centre of the cortex, whereas the second region is positioned close to the outer edge of the cortex.
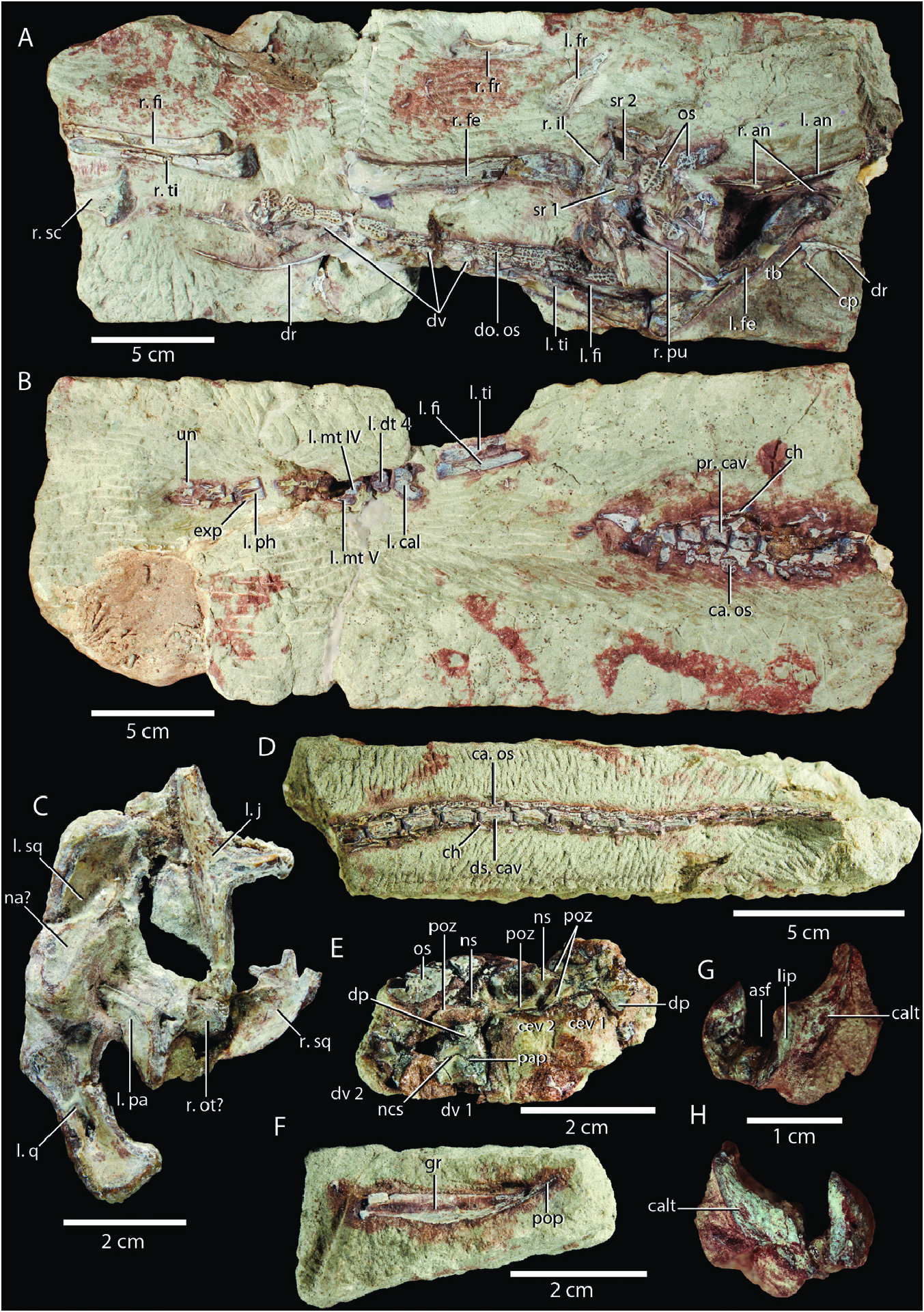

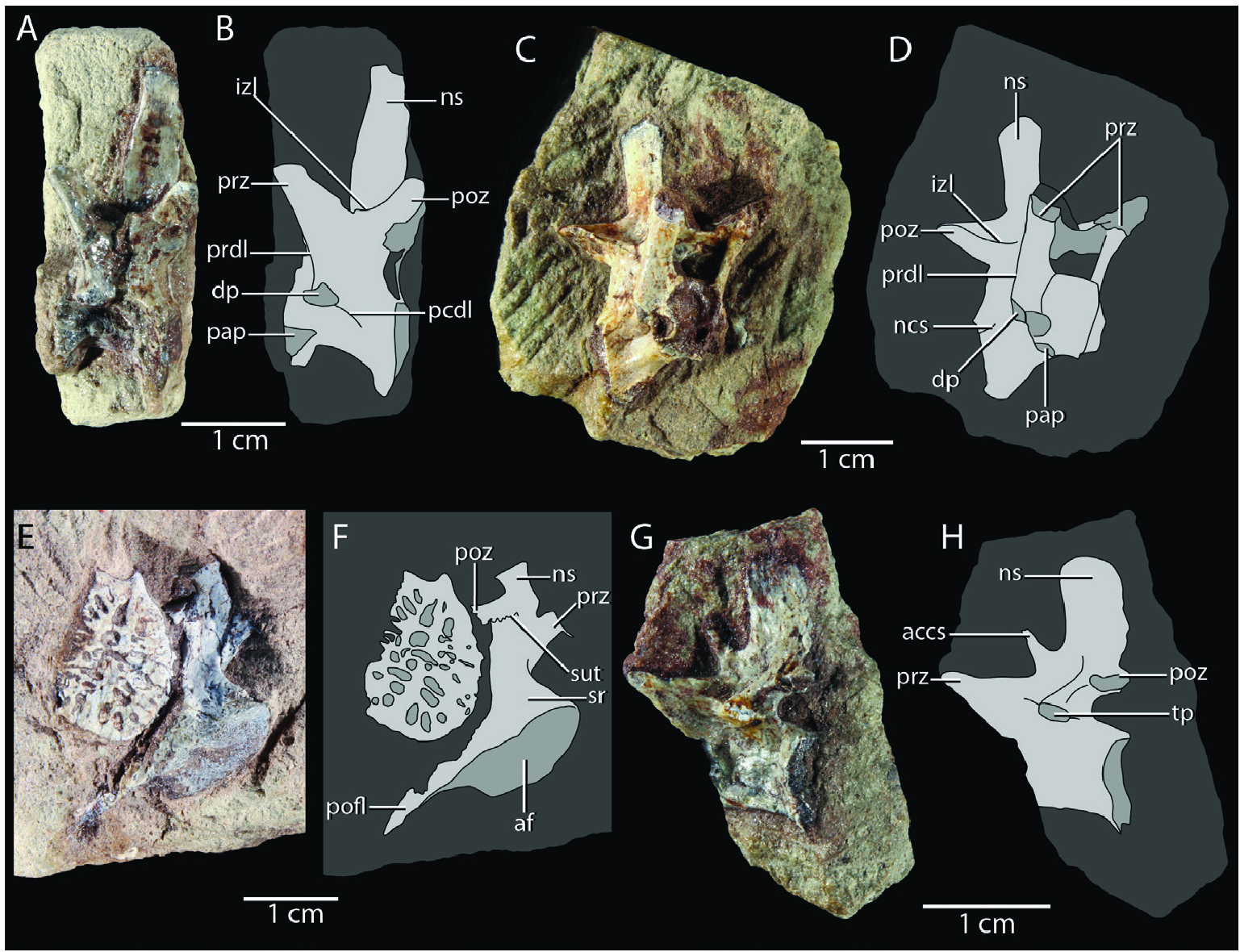

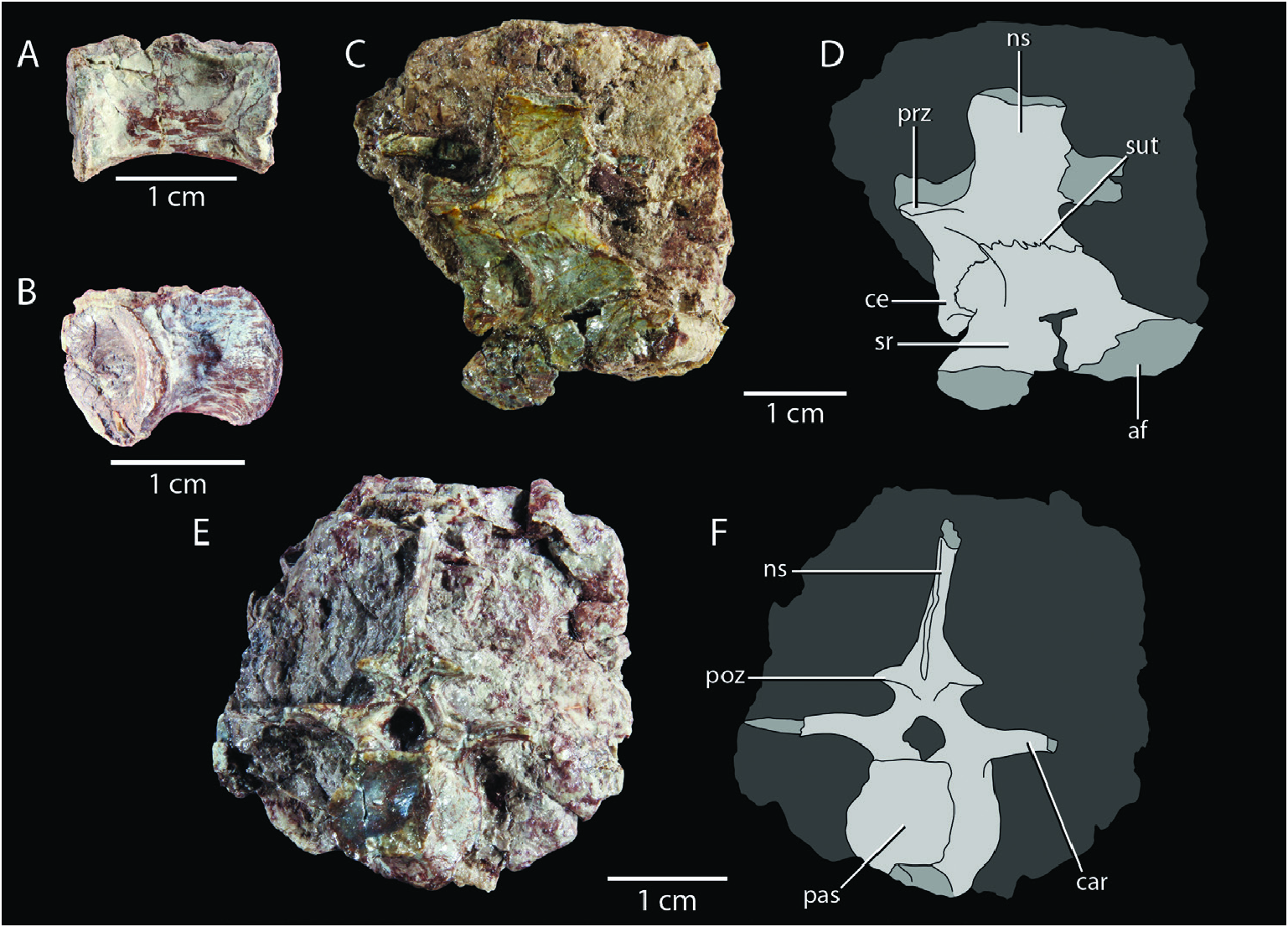
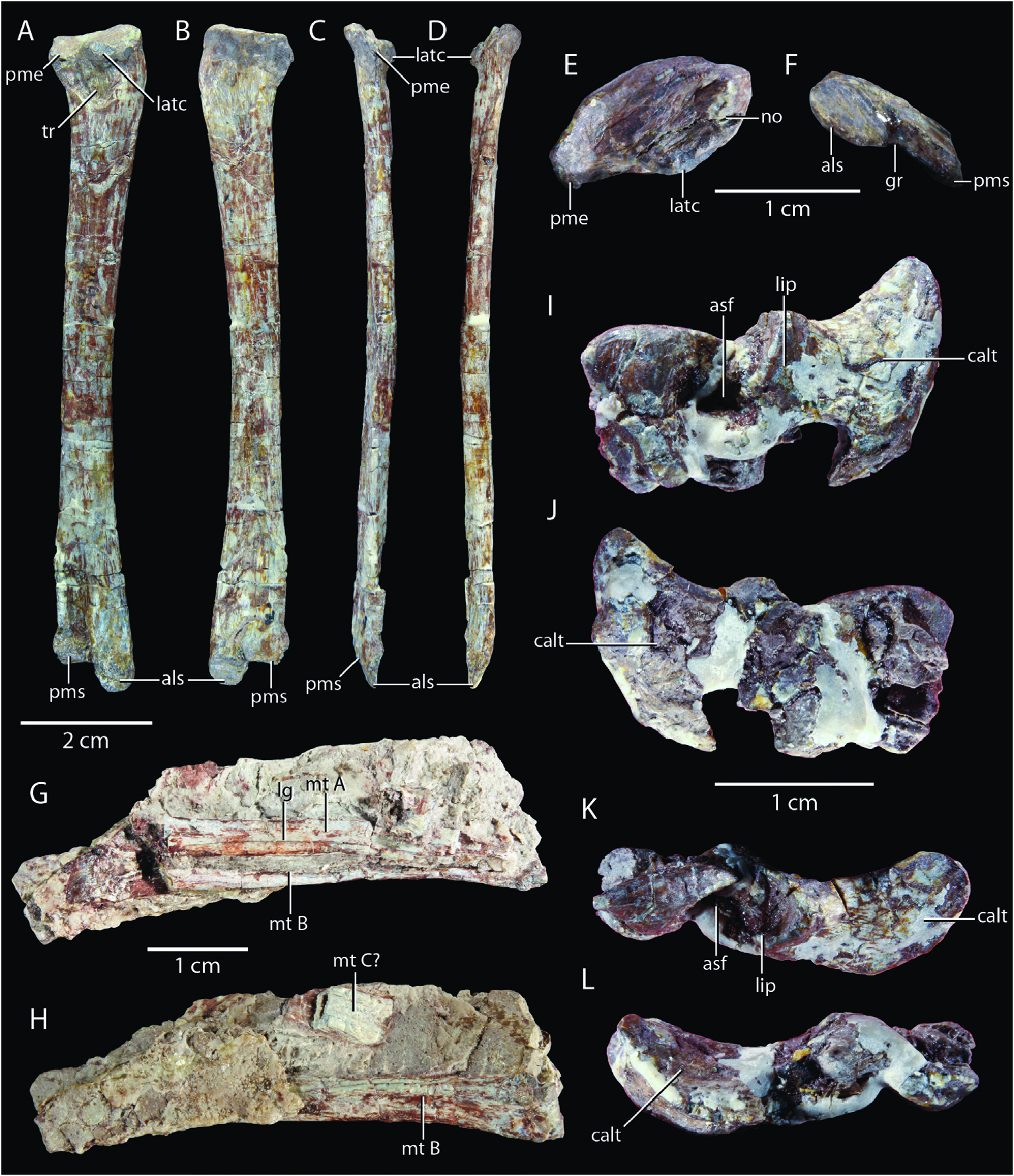


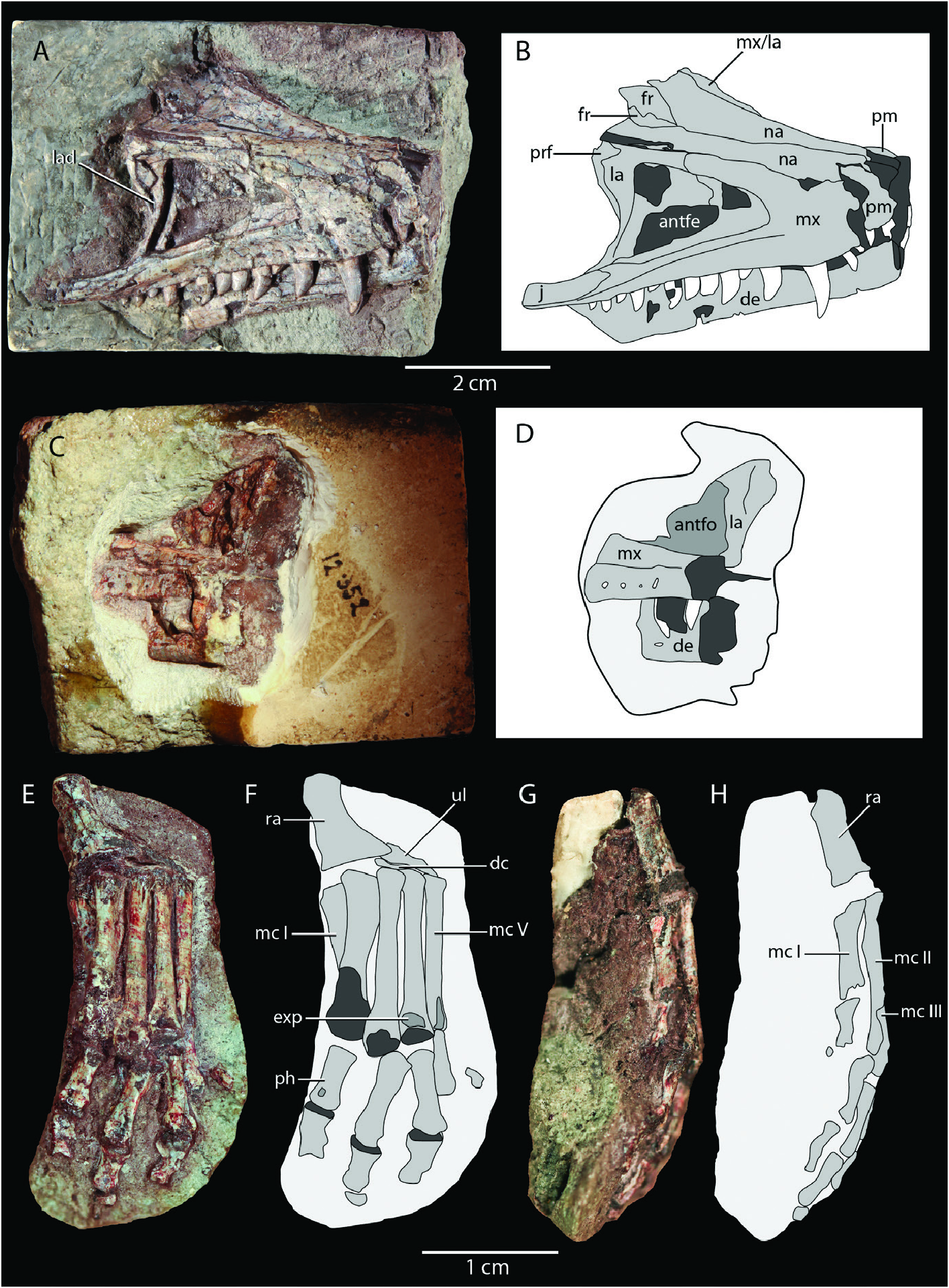
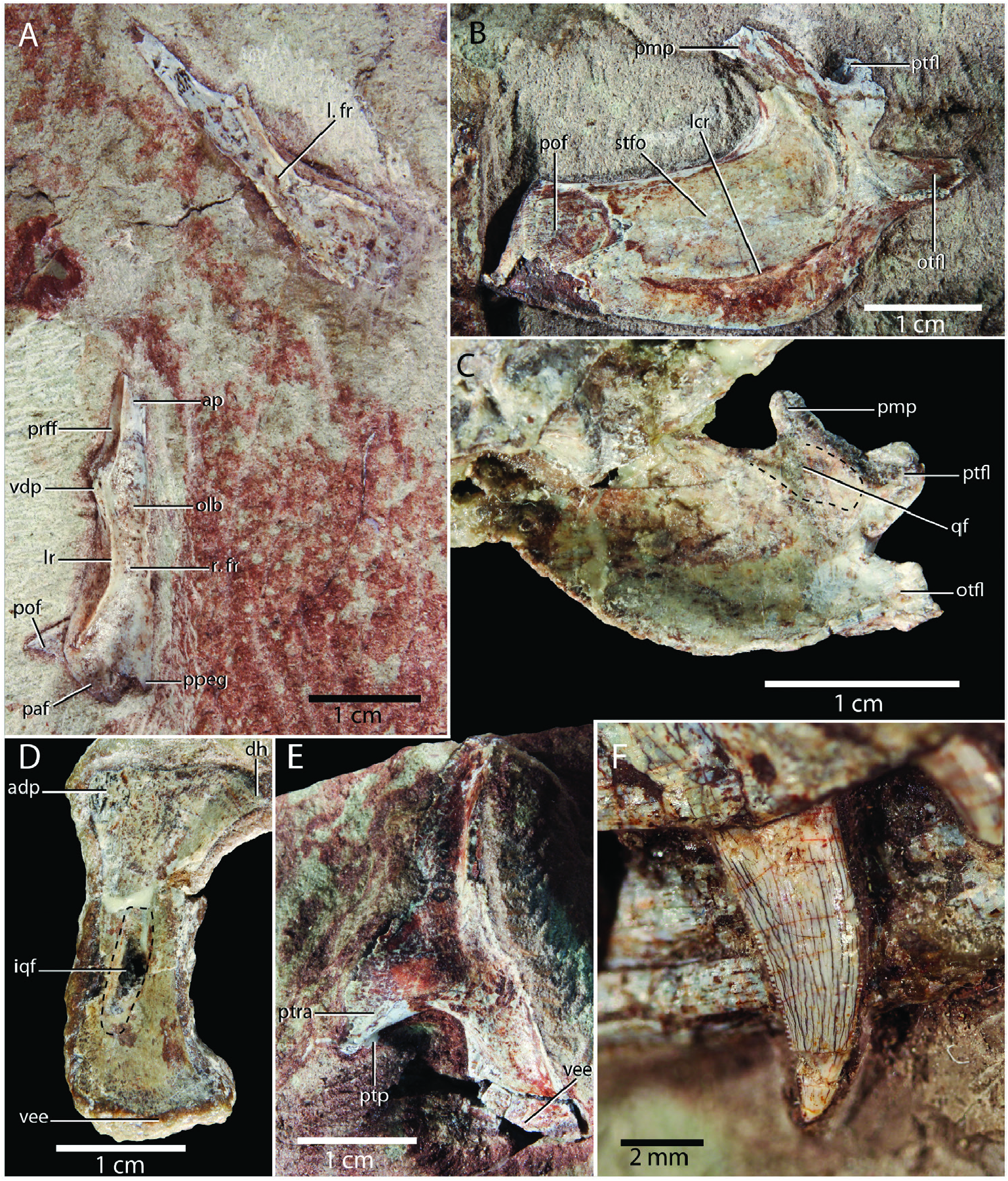

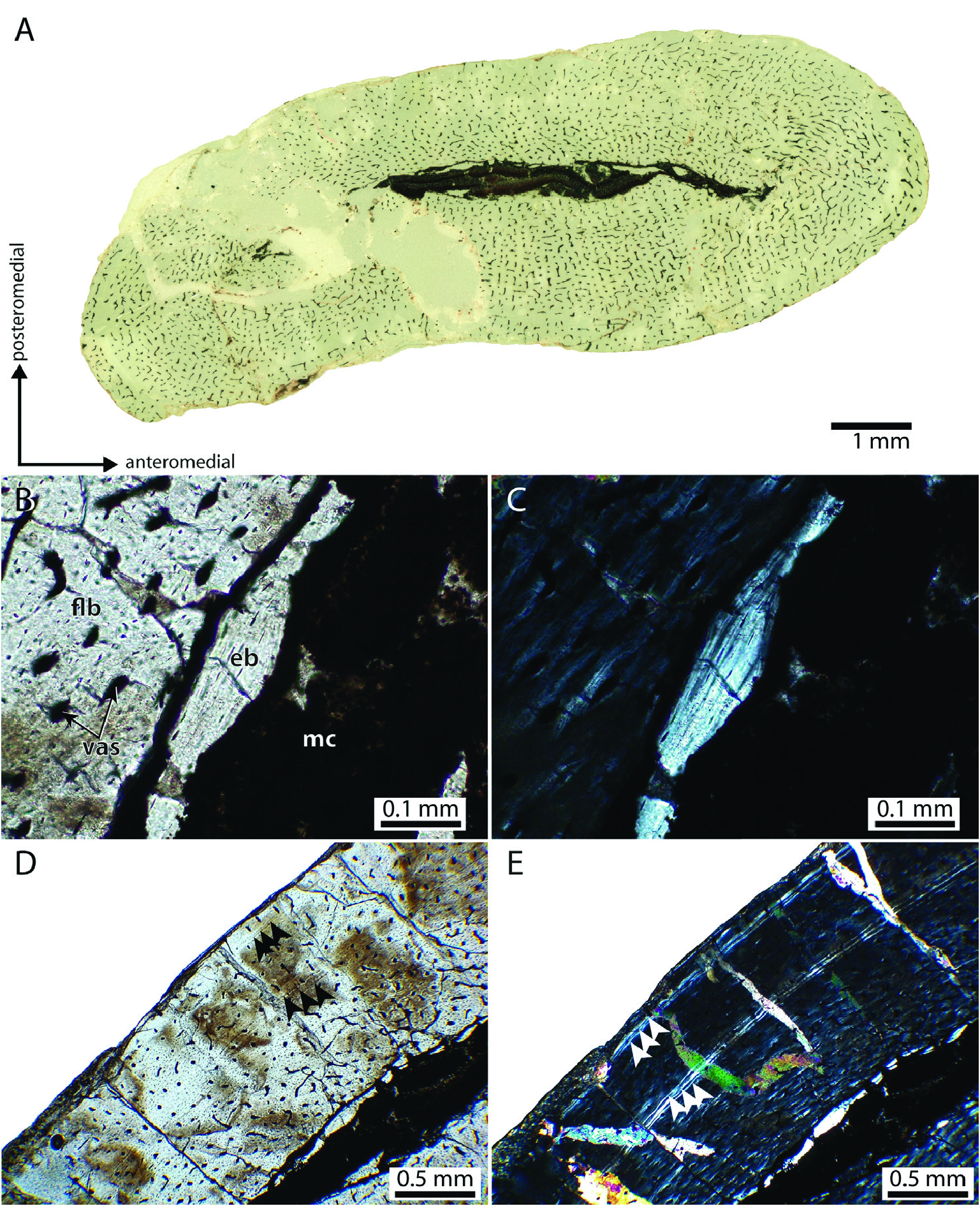
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.