
Eremiascincus phantasmus, Mecke, Sven, Doughty, Paul & Donnellan, Stephen C., 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3701.5.1 |
|
publication LSID |
lsid:zoobank.org:pub:26CA85BD-BA77-411C-9D4D-9C4706BB8F1A |
|
DOI |
https://doi.org/10.5281/zenodo.5613170 |
|
persistent identifier |
https://treatment.plazi.org/id/03A387CC-FF82-FFE8-FF7C-F985FE377776 |
|
treatment provided by |
Plazi |
|
scientific name |
Eremiascincus phantasmus |
| status |
sp. nov. |
Eremiascincus phantasmus sp. nov.
English name: Ghost Skink
German names: Südlicher Streifenskink, Gespenstskink ( Figs. 13 View FIGURE 13 & 14 View FIGURE 14 )
Synonyms:
Sphenomorphus fasciolatus (part.) Storr, 1974 Rec. West. Aust. Mus. 3 (1): 68–70 (‘Ghost Skink’ = p. 69) Eremiascincus fasciolatus (part.) Greer, 1979; Rec. Aust. Mus. (32) 7: 323 et seqq. (‘Ghost Skink’ = p. 325) Eremiascincus fasciolatus (part.) Cogger, 2000; Rept. Amphib. Aust.: 477
Type material of the species herein described as new has been deposited in the SAMA, QM and AMS collections.
Holophoront: SAMA R49358 (field number C97047 View Materials ) ( Fig. 13 View FIGURE 13 ), male. Type locality: collected 5.5 km SE of Red Lake Yard, E side of Lake Hope Channel, SA (28°15'29''S, 139°12'29'' E); leg. M. Hutchinson.
Paratypes (22 specimens): SAMA R49896 (F)— 43.7 km east of Purni Bore, on the Rig Road, SA, 26°19'17''S, 136°31'53''E; R50047 (unsexed juvenile)— 66.8 km E Purni Bore, Big RD, SA, 26°19'17''S, 136°31'53''E; R50888 (F)— 29.1 km NNW Murda Hill, Simpson Desert Regional Reserve, SA, 26°42'09''S, 138°15'44''E; R51482 (M)— 25.1 km NW Atna Hill, Simpson Desert CP, SA, 26°16'29''S, 137°19'37''E; R51436 (M)— 1.1 km SSE Approdinna Attora Knolls, SA, 26°04'50''S, 137°36'43''E; R54137 (M)— 37.1 km WNW of Muloorina Homestead, SA, 29°09'45''S, 137°32'01''E; R54255 (unsexed juvenile)— 15.3 km WNW Kannakaninna Waterhole, Kalamurina, SA, 27°53'09''S, 137°49'13''E; R55437 (F)— 17 km NW Karrathunka Waterhole, SA, 26°06'07''S, 139°08'45''E; R55447 (M)— 12.4 km NW Karrathunka Waterhole, SA, 26°08'27''S, 139°09'54''E; R55507 (M)— 30.2 km ESE Mulka Hill, SA, 28°26'51''S, 138°52'06''E; AMS R 113164 (M)— 25 km N of Poeppel Corner, Simpson Desert, NT, 25°46' S, 138°00'E; R155329 (M)—Sturt National Park, 1.8 km W (by road) along Whitecatch Gate Road, NSW, 29°06'51''S, 141°10'54''E; QM J41600 View Materials (F), J41602 View Materials (F)—Durrie Station, Glennie Well, QLD, 25°55'48''S, 139°55'49''E; J44070 View Materials (F)—Simpson Desert, QLD, 25°40'48''S, 138°31'48''E; 48473 (M)— Chookoo, 30 km WSW Jackson, QLD, 27°34'48''S, 141°54'00''E; J75056 View Materials –57 (M), J75060 View Materials (M), J75067 View Materials (unsexed juvenile)— 2 km W of Birdsville (behind Airstrip), QLD, 24°53'60''S, 139°18'00''E.
Diagnosis and Description based on material cited in the Appendix and color photographs by P. Tremul (see also Fig. 14 View FIGURE 14 ).
Taxonomic history. Whereas Waite’s (1929) concept of Hinulia fasciolata in southern Australia is rather unsatisfactory and misleading as the morphology for the SA populations seems to be largely based on the type description in Boulenger (1887) (see also Greer [1979] for a discussion on data of reproduction for Hinulia fasciolata presented by Waite [1929]), Worrell (1963) was probably the first author who used the colloquial name ‘Ghost Skink’ for a pale, undescribed species of desert-dwelling skink and attributed the scientific name Lygosoma (Sphenomorphus) monotropis to that species. Lygosoma (Sphenomorphus) monotropis , however, is a junior synonym of the heavily banded Eremiascincus richardsonii (Cogger et al. 1983) and is clearly distinct from the form Worrell described. The area of distribution Worrell suggested includes the central parts of SA and the arid parts of WA. Because Worrell did not include ‘ pallidu s’ in his Reptiles of Australia it may be that he believed the ‘Ghost Skink’ and ‘ pallidu s’ to be conspecific. Although Worrell’s species concept remains mysterious, and Lygosoma (Sphenomorphus) monotropis sensu Worrell (1963) might be better regarded a nomen ambiguum rather than a synonym, the name ‘Ghost Skink’ was generally used by subsequent authors (e.g., Storr 1974; Greer 1979) to refer to the patternless, extreme pale form of " E. fasciolatus " from the Lake Eyre region of north-eastern SA.
Diagnosis. A medium to large-sized (snout-vent length up to 92.5 mm), slender narrow-banded Eremiascincus having 8–9 undivided supralabials (usually 8); 2 infralabials usually in broad contact with the postmental scale; 20–30 subdigital lamellae, at least basally in two rows and at least basally keeled; plantar scales 12–18, smooth; 28–33 scale rows at mid-body; head small, snout depressed, and ear opening small and circular; dorsum with keels posteriorly and tail often with strong ridges; coloration pattern usually comprises 29–39 perfectly transverse narrow, often very pale bands on the tail (type a, b); body without bands or with indication of light narrow bands (type a, b) dorsally or more frequently on sides of body, usually pale, without any indication of banding visible.
Description. Dimensions and general characteristics. A medium to large-sized (snout-vent length of adult specimens 62.6–92.5 mm, mean 81.2, n = 117), slender Eremiascincus ; head small (HeadL1/SVL 0.17–0.20, mean 0.18, n = 113); snout long (SnoutL/HeadL1 0.39–0.46, mean 0.42, n = 113) top of snout depressed, tip of snout obtusely pointed; canthus rostralis rounded; lower eyelid movable, scaly; ear opening circular in shape and small (maximum length 1.8 mm, maximum height 1.8 mm), about a quarter the area of eye aperture, with minute granules at anterior border; ear opening followed by a depression of the integument; tympanum sunk; limbs pentadactyl, overlapping when adpressed, moderately long (hind legs ~40% length of snout-vent length); fingers and toes moderately long, circular in section; third and fourth finger length similar; fourth toe longer than third; tail up to ~138% of SVL (range 122.7–137.8%, mean 129.1%, n = 15), with strong ridges or keels, round in cross section; hemipenis deeply bifurcated, not columnar.
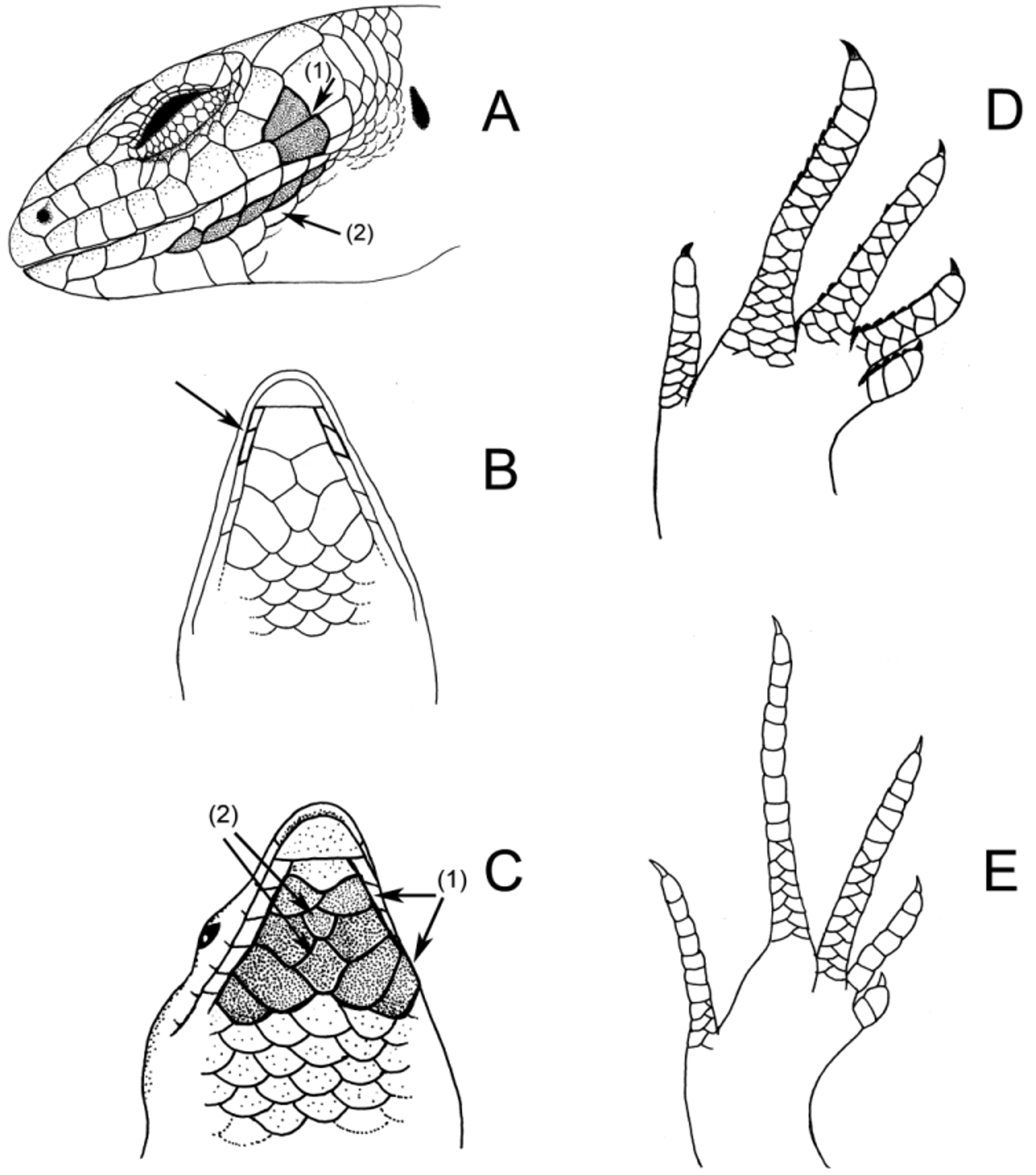
Scalation. Nasals widely separated; frontal arrowhead-shaped, ~70% longer than interparietal, much larger than prefrontals; width of rostral-frontonasal suture less than half width of frontonasal; prefrontals narrowly separated; frontal contacting frontonasal; frontoparietals free (paired); interparietal normally free, almost as large as a frontoparietal; parietals large, in contact behind interparietal; each parietal bordered posteriorly by upper secondary temporal and (if present) enlarged nuchal scales; nuchals 0–1; supraoculars 4, normal in shape and orientation; medial 2 much longer than wide, 2 or 3 in contact with frontal; loreals 2, second usually wider than tall, squarish or rectangular; supranasal scale absent; preoculars 2, lower larger; presuboculars usually 2; supraciliaries 7–10 (mostly 8, mean 8.4, n = 117), first largest; supralabials 7–9 (usually 8, mean 8.0, n = 117), supralabial scales undivided, last supralabial scale overlaps lower secondary temporal; infralabials 6–9; postmental in contact with first and second infralabial ( Fig. 17 View FIGURE 17 B); no trace of subinfralabial scales; 1 primary and 2 secondary temporals; 28– 33 (mostly 30–32) scales at mid-body (mean 30.3, n = 113), laterals smallest; 60–71 paravertebral scales (mean 65.9, n = 114); 20–30 subdigital lamellae under fourth toe (mean 25.2, n = 114), scale under fourth toe divided medially, at least basally but often grooved for more than half of digit; weakly keeled or callose (with a single keel per side of a grooved scale); scales on top of toes variable, usually with multiple series covering base or up to half of the length of the digits, sometimes only single scale rows present; scales on soles of hands and feet granular and smooth; body and head shiny-scaled; scales on body smooth; trailing edges of dorsal and lateral scales angularly three-sided rather than smoothly rounded; subcaudals larger than ventrals; tail above usually strongly keeled (keels forming ridges); 2 median precloacal scales distinctly enlarged.
Coloration and pattern in preservative (ethanol). Upper surface (head, dorsum, tail, and limbs) whitish to pale gray, rarely with a fine banding discernible laterally or dorsolaterally (never forming conspicuous dark bands across the dorsum); bands usually absent altogether or only evident on the tail as pale brown cross-bands of a single scale row or less and separated by 2 rows of paler scales (if discernible 29–39 bands on tail) (type a, b), regenerated tails usually display no banding; subocular region appears to be dark bluish because of the underlying eye socket; labials pale; underneath immaculate ivory or silvery white, sometimes displaying a strong contrast between the pale dorsal and the ivory or white lateroventral side; scales on chin and gular region, palmar and plantar scales not colored differently; iris as dark as pupil; tongue pale. Hatchlings are similar in most respects to adults but with more intense bands on the tail, which become more faded with age.
Coloration and pattern in life. Ground color pale yellowish, brownish or whitish with very faded caudal striping; sometimes with obscure indication of bands laterally or dorsolaterally, rarely with a fine very light banding on dorsum; sides are usually paler than dorsum; belly cream to pinkish; all body scales glossy. Juveniles sometimes display a maculated dorsum. The coloration has resulted in the common name ‘Ghost Skink’. Fig. 14 View FIGURE 14 shows a specimen in life.
Craniology. According to the data presented in Greer (1979), this species has 9 premaxillary teeth, the surface exposure of the postorbital bone in the supratemporal arch is short, and an ectopterygoid process is not evident or only weakly developed.
Sex ratio and dimorphism. The sex ratio of specimens favored males (67:50). All meristic and morphometric variables were tested for sexual dimorphism, with significant differences found for ArmL (males: mean = 21.4, SD = 1.3, median = 21.6, n = 66; females: mean = 20.5, SD = 1.2, median = 20.6, n = 49; Z = -3.718***), LegL (males: mean = 30.3, SD = 2.0, median = 30.3, n = 66; females: mean = 28.8, SD = 1.7, median = 28.8, n = 49; Z = -4.215***), FootL (males: mean = 12.1, SD = 0.9, median = 12.2, n = 66; females: mean = 11.6, SD = 0.7, median = 11.5, n = 49; Z = -3.357***), ToeL3 (males: mean = 6.2, SD = 0.5, median = 6.3 n = 65; females: mean = 5.9, SD = 0.5, median = 5.9 n = 48; Z = -3.177**), ToeL4 (males: mean = 8.4, SD = 0.6 median = 8.4, n = 66; females: mean = 8.1, SD = 0.6, median = 8.0, n = 49; Z = -2.883**), HeadL1 (males: mean = 15.4, SD = 1.1, median = 15.7, n = 64; females: mean = 14.3, SD = 1.0, median = 14.4, n = 49; Z = -4.588***), HeadL2 (males: mean = 14.1, SD = 1.0, median =14.4, n = 65; females: mean = 13.3, SD = 1.0, median = 13.5, n = 50; Z = -4.203***), HeadW (males: mean = 9.5, SD = 0.8, median = 9.6, n = 64; females: mean = 8.8, SD = 0.8, median = 8.8, n = 49; Z = -4.372***), SnoutL (males: mean = 6.4, SD = 0.5, median = 6.5, n = 67; females: mean = 6.0, SD = 0.4, median = 6.0, n = 50; Z = -4.243***), HeadH (males: mean = 6.6, SD = 0.6, median = 6.7, n = 65; females: mean = 6.1, SD = 0.5, median = 6.2, n = 47; Z = -4.584***). The following ratio likewise showed significant differences between sexes: HeadL1/SVL (males: mean = 0.19, SD = 0.00, median = 0.19, n = 64; females: mean = 0.18, SD = 0.01, median = 0.18, n = 49; Z = -5.182***). TrunkL/SVL did not differ between sexes, but probability level was close to the critical level.
Details of holophoront ( Fig. 13 View FIGURE 13 ) (SAMA R49358) [adult male]. SVL 90.5 mm, TrunkL 50.6 mm, TailL 109.1 mm (tip regenerated), ArmL 21.9 mm, LegL 29.9 mm, HeadL1 15.8 mm, HeadL2 14.9 mm, HeadW 10.3 mm, SnoutL 6.7 mm, snout depressed, HeadH 6.6 mm, AxillaEar 16.9 mm, ear opening 1.3 x 1.3 mm, MBSR 28, PVS 65, SupraLab 9 (DividedLab: none/undivided), InfraLab 8, SupraCil 9, prefrontals separated, presuboculars 2, nuchals 1, 4TLam 25, upper surface of fourth toe with scales in single rows with transverse sutures along almost entire digit, plantar scales 13, no bands or pattern noticeable on body, BandsTail (type b) 32.
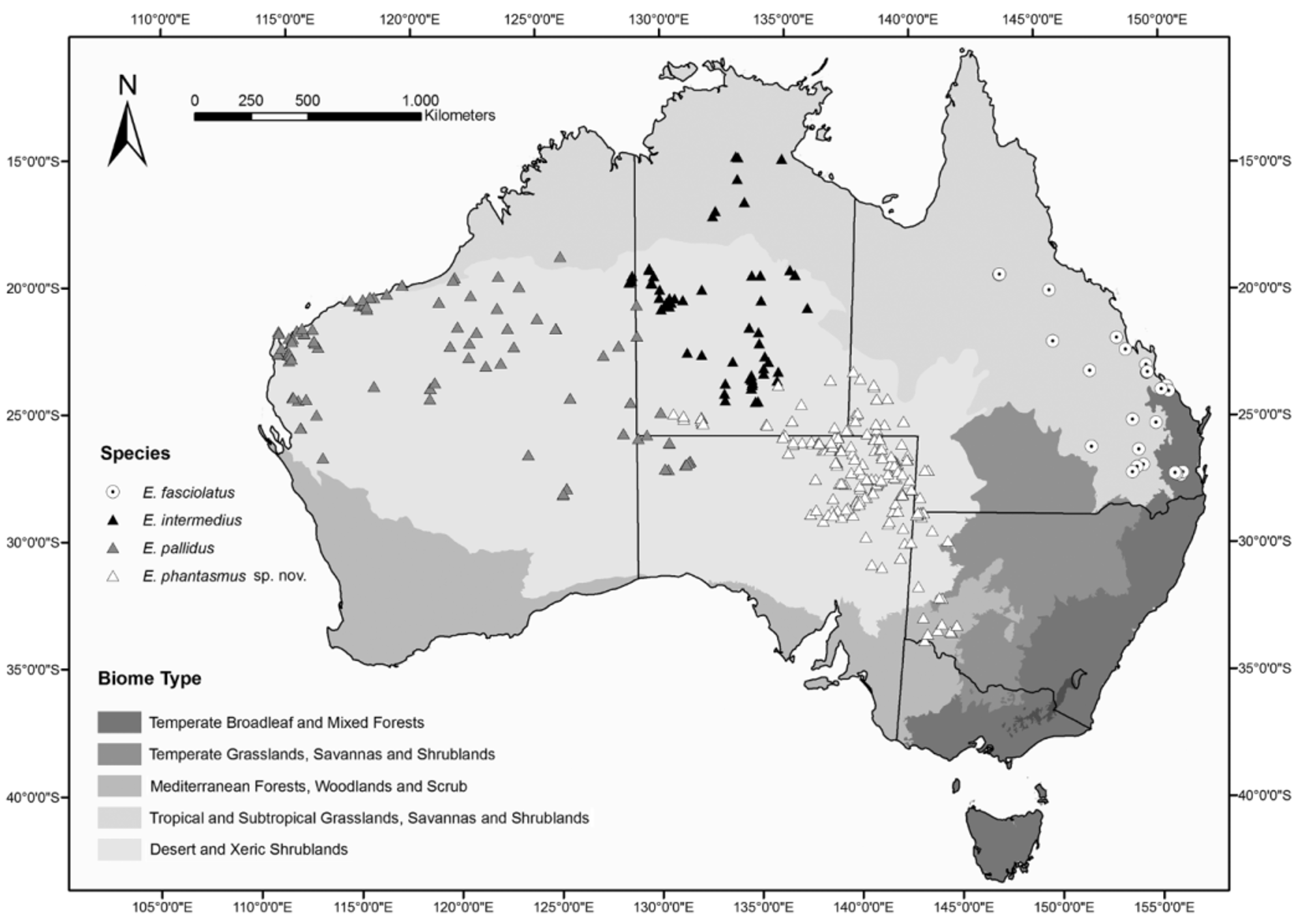
Distribution. Eremiascincus phantasmus sp. nov. predominantly occurs in the low elevation deserts of the Lake Eyre Basin with the main distribution area in north-eastern SA ( Fig. 18 View FIGURE 18 ). The area of distribution extends from south of Lake Frome north-west to the mound spring area of Emerald and Strangways Springs and north to Witjiara National Park at the western border of the Simpson Desert, where it predominantly occurs in the Marla- Oodnadatta and the Marree Soil Conservation Districts. In the NT, E. phantasmus sp. nov. extends from Andado in the south, north-west to Charlotte Waters with the northernmost records from Illogwa Creek in the south-east central NT and Lake Caroline in the east. In the south-eastern corner of the NT, E. phantasmus sp. nov. is poorly collected. The species also occurs in the Central Ranges xeric scrubland in the Curtin Spring and Uluru area, where it may be sympatric with E. pallidus . Eremiascincus phantasmus sp. nov. has not been collected from the central Finke bioregion. In QLD the species occurs in the Channel Country of the extreme south-west. It occurs from the Mirrica Bore region in the north, south to Mount Leonard. The species is also found in the Baryulah area south to the border of NSW. From Cameron’s Corner, the species extends south-east to Tero Creek. In south-west NSW the distribution extends from the Broken Hill area in the north to Wentworth on the Victorian border and east to Mungo National Park (see also Swan 1990 for a report of an eastern range extension). Eremiascinus phantasmus sp. nov. to date has not been recorded from the far north-west of Victoria.
Geographic variation. Variation in color pattern (e.g., subdued narrow dorsolateral bands) is present in some specimens, but in other respects (e.g., labial scalation) these localized entities are typical of E. phantasmus sp. nov.
Habitat and ecology. The species inhabits low lying flood-prone areas and sandy rises with Spinifex cover or without vegetation ( Fig. 15 View FIGURE 15 ) and is often found in association with buildings and under rubbish piles. The very glossy, smooth-scaled skink is a sand-swimmer as one would expect from its overall morphology and habitat and occurs on substrates sandy enough to support a range of burrowing species (P. Tremul, pers. comm.). Eremiascincus phantasmus sp. nov. is crepuscular and nocturnal in its activity, becoming most active in the late afternoon and the first few hours of darkness. Like its congeners, the species feeds on insects, small skinks, and geckos. Clutch size varies from 2– 7 eggs, measuring approximately 16–17 x 10 –12 mm. Hatchlings have a SVL of 32–36 mm and a TailL of 72–75 mm (D. Brown, pers. obs. of E. phantasmus from QLD held in captivity).
Etymology. The specific name ‘ phantasmus ’ is Latin for ‘ghost’ and used as a noun in apposition. It is the Latin translation for a vernacular name Worrell proposed in 1963.
Comparison with other species. This medium-sized to large Eremiascincus is generally distinguishable from all other narrow-banded skinks by its pale dorsal coloration. This character is only shared with the smaller E. pallidus from the western arid zone, which is distinguishable from E. phantasmus in usually having 7 supralabial shields (instead of usually 8) and 1 infralabial in contact with the postmental scale (vs. almost always 2 infralabials in contact with the postmental).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.