
Potamon fluviatile (Herbst, 1785)
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2009.00625.x |
|
persistent identifier |
https://treatment.plazi.org/id/03A387D4-1B13-FFC2-53DA-F8A10EEA5B12 |
|
treatment provided by |
Valdenar |
|
scientific name |
Potamon fluviatile |
| status |
|
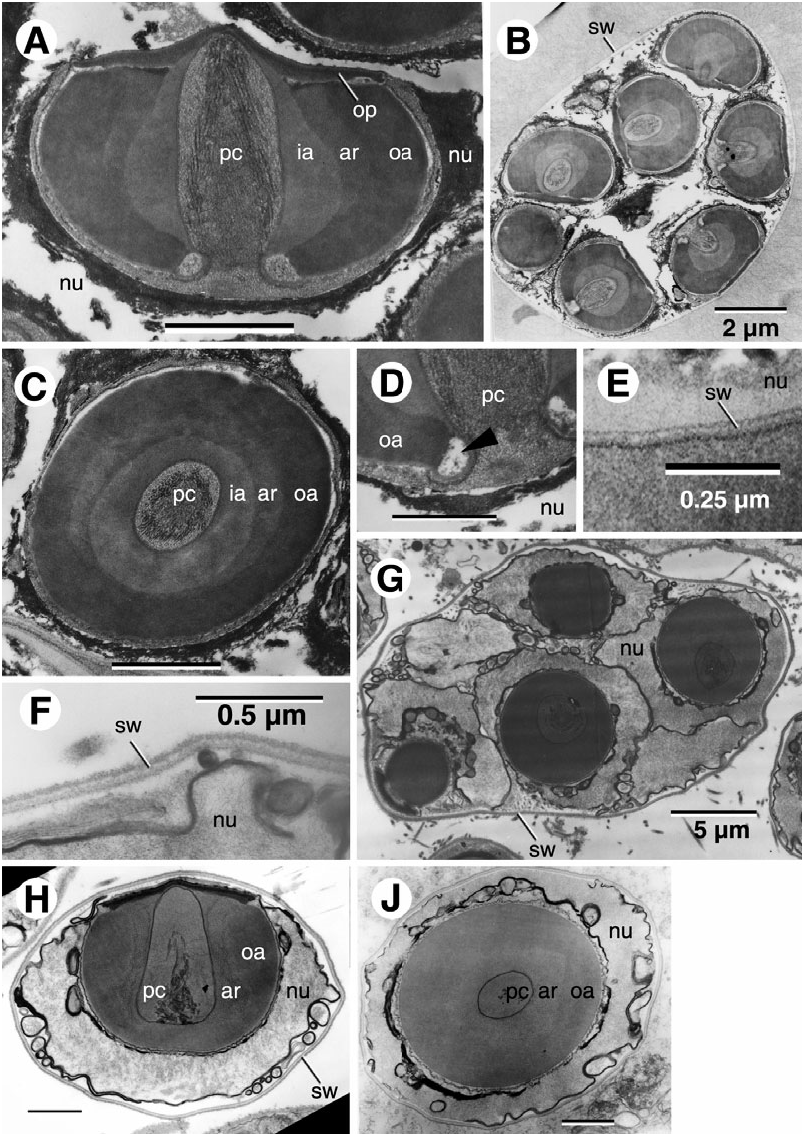
POTAMON FLUVIATILE View in CoL ( FIG. 1F–J; TABLE View Figure 1 2)
As stated above, this specimen of Potamon fluviatile was frozen after death, which elicited acrosome reaction. However, there were still a few sperm cells that remained intact. The operculum is rather thin. Lateral of the operculum there are electron-lucent structures that resemble a vestigial periopercular rim as it occurs in Potamonautidae and Gecarcinucidae . The acrosome ray zone is prominent and bulging outwards. There are many laterally situated nuclear arms. Potamon fluviatile shows both coenospermia and cleistospermia, with the coenospermic spermatophores being identical to those of the Potamiscinae .
PHYLOGENY OF THE POTAMIDAE
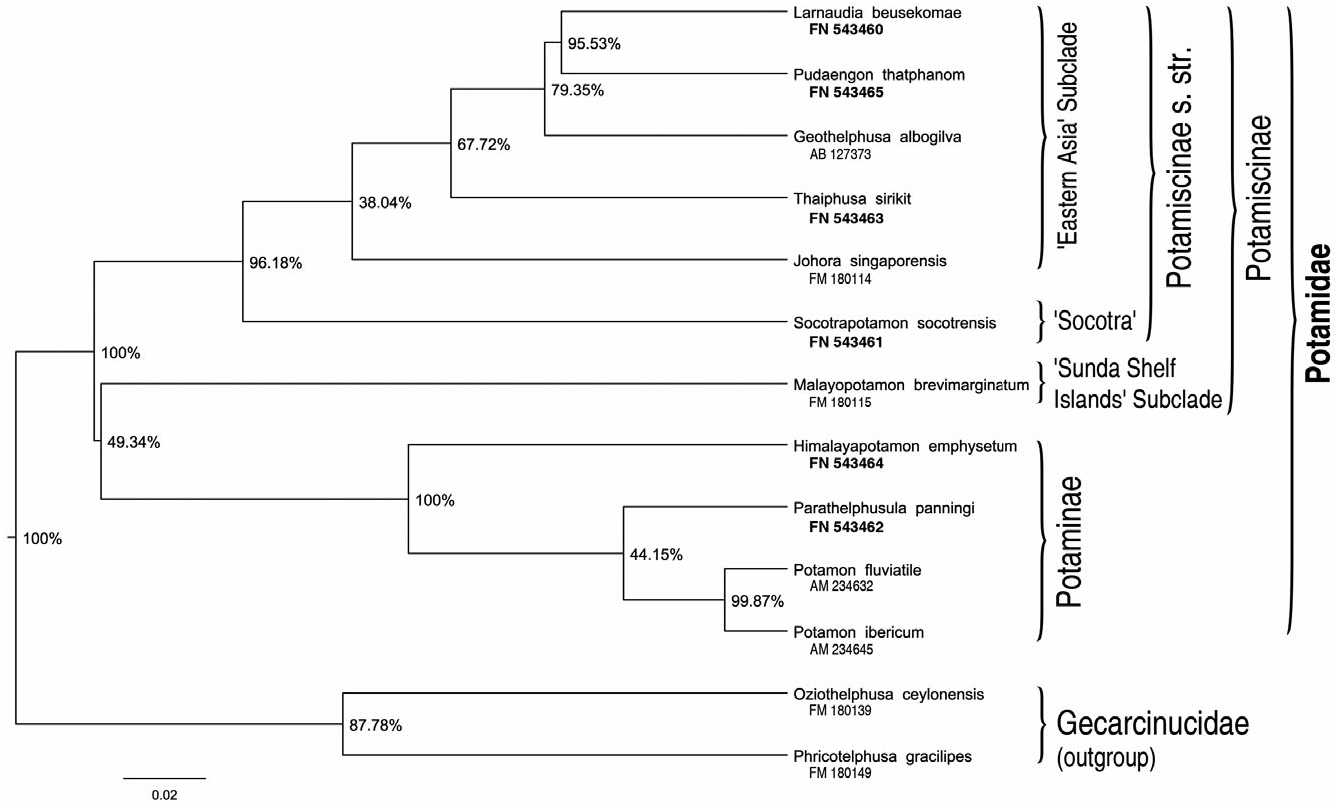
Here, phylogenetic inference is not performed to reveal potamid relationships, but to be used in the subsequent ancestral character state reconstruction. Thus, it is important to note that the topology of the 16S rDNA analysis (although several splits have only minor support; Fig. 2 View Figure 2 ) is widely in agreement with that of Shih et al. (2009), which was based on a large data set of 72 species and 49 genera. The major difference is the position of Malayopotamon brevimarginatum (representing the ‘Sunda Shelf islands’ subclade), which clusters in our analysis with a very short branch and low posterior probability as sister group to the subfamily Potaminae , whereas in the analysis of Shih et al. (2009) it is monophyletic with the other Potamiscinae (however, only weakly supported in their minimum-evolution approach). Apparently the ‘Sunda Shelf islands’ subclade corresponds to the Isolapotamidae of Bott (1970a), and might be recognized as a third potamid subfamily Isolapotami-
nae (see Shih et al., 2009). We did not force monophyly on the Potamiscinae in the phylogenetic analysis, but kept this incongruence during the ancestral state reconstructions as none of the two topolo-
gies can currently be excluded. In summary, we can recognize the ‘ Potamiscinae s.s.’ (including the genus
Socotrapotamon , but excluding Malayopotamon ), the subfamily Potaminae , and a group [ Malayopotamon
+ Potaminae ], with the possibility that the latter is not monophyletic (the respective node is present in
49% of the 1000 trees fed into the ancestral character state analysis).
ANCESTRAL CHARACTER STATE RECONSTRUCTION
Reconstructions of ancestral character states under a parsimony model are shown in Figure 5A–D. The ancestral character state of the operculum (perforate or imperforate) at the base of the Potamidae remains equivocal. In total, 25.2% of reconstructions support the imperforate state and 19.5% of reconstructions support the perforate state at this node (Fig. 5C). The same ambiguous reconstruction of operculum states occurs in the ‘ Potamiscinae s.s. ’, the Potaminae , and the [ Potaminae + Malayopotamon ].
A true middle acrosomal zone only occurs in the
Potamiscinae and in the genus Malayopotamon
(Fig. 5D). Consequently, the absence of the middle acrosomal zone belongs to the ground pattern of the
Potaminae . However, because of its absence in
Socotrapotamon , the ancestral state reconstruction finds this character in only 58% of the analysed trees at the base of the ‘ Potamiscinae s.s.’.
The presence of the ‘tongue-and-groove’ connection that connects the operculum and acrosomal zones situated beneath is most probably an apomorphy for the whole Potamidae (occuring in 71.1% of
·
Figure 5. Reconstruction of ancestral sperm characters under a parsimony model over 1000 trees from the Bayesian inference (MESQUITE 2.6). A, operculum perforate or imperforate. B, ‘middle acrosome zone’ present or absent.
C, ‘tongue-and-groove’ connection present or absent. D,
coenospermia present or absent. Values at nodes indicate the percentage of trees with the respective state reconstructed or percentage of equivocal reconstructions. The proportion of trees that failed to find a respective node is the same for all four reconstructions, and the exact values are given in Figure 5B.
reconstructions; Fig. 5A). Secondary reduction of this character took place in Socotrapotamon socotrense , and in the lineage leading to Parathelphusula panningi and the genus Potamon within the Potaminae .
The coenospermic spermatophores are reconstructed as the ground pattern of the Potamidae (reconstructed in 80.3% of the trees analysed; Fig. 5B). Several reductive events led to cleistospermia in the genus Potamon , in Johora singaporensis , and in Thaiphusa sirikit . Also for the ‘ Potamiscinae s.s. ’ (79.1%), the [ Potaminae + Malayopotamon ] (40.0%), and for the Potaminae (81.7%), coenospermia most probably represents the ancestral character state.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |