
Adelophryne patamona, Macculloch, Ross D., Lathrop, Amy, Kok, Philippe J. R., Minter, Leslie R., Khan, Samir Z. & Barrio-Amorós, César L., 2008
|
publication ID |
https://doi.org/ 10.5281/zenodo.274492 |
|
DOI |
https://doi.org/10.5281/zenodo.5613154 |
|
persistent identifier |
https://treatment.plazi.org/id/03A3E47E-FFF2-7946-FAA1-B2D10C4CE295 |
|
treatment provided by |
Plazi |
|
scientific name |
Adelophryne patamona |
| status |
sp. nov. |
Adelophryne patamona View in CoL , sp. nov.
Figs 1–7 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7
Material. Holotype. ROM 43034, an adult male, from the north slope of Mount Wokomung, Guyana, 05º 06' 35" N, 059º 49' 15" W, 1234 m, collected by A. Lathrop, S. Khan and R. MacCulloch, 1 November 2004.
Paratypes. ROM 43035, an adult female, from the north slope of Mount Wokomung, Guyana, 05º 05' 33" N, 059º 50' 35" W, 1411 m, collected by A. Lathrop, S. Khan and R. MacCulloch, 7 November 2004; IRSNB 1971–4, adult females, from the east base of Mount Maringma, Guyana, 05º 12' 38" N, 060º 33' 59" W, 1060 m, collected by P. Kok, P. Benjamin and C. Perry, 20 November 2007; IRSNB 1975, an adult female, IRSNB 1976-7, two juveniles and IRSNB 1978, an adult male from the southeast slope of Mount Maringma, Guyana, 05º 12' 16" N, 060º 34' 39" W, 1376 m, collected by P. Kok, P. Benjamin and C. Perry, 22–24 November 2007.
Etymology. The specific epithet is a noun in apposition, and refers to the Patamona people, who live in the area where the holotype was collected.
Diagnosis. A large species of Adelophryne , characterised by maximum SVL> 20 mm, second phalange of Finger IV longer than wide, tips of digits with small pointed discs, tympanum small, separated from eye by a distance equal to tympanum diameter, posterodorsal part of tympanum covered by a fold, and an inconspicuous glandular ridge from tympanum to arm insertion.
The specimens can be placed in the genus Adelophryne because they possess the following combination of characters: small size, flattened digits with indistinct subdigital pads rather than subarticular tubercles, shortened Finger IV with single subdigital pad, discs with pointed tips and lateral grooves, vomerine teeth present, tongue narrow, tarsal tubercle absent.
Of the other genera of small terrestrial frogs in the region, Noblella has a tarsal tubercle and no vomerine teeth, Phyzelaphryne has a broad tongue, round digits, and discs with circumferential grooves, and Pristimantis has two subarticular tubercles on Finger IV. The differences among these genera were elaborated upon by Hoogmoed and Lescure (1984).
Adelophryne patamona can be distinguished from A. adiastola and A. pachydactyla by having 3 phalanges in Finger IV (2 phalanges in A. adiastola and A. pachydactyla ) and large size, up to 23 mm SVL (largest A. adiastola 14 mm, largest A. pachydactyla 11.1 mm).
Adelophryne patamona can be distinguished from A. baturitensis by the presence of subdigital pads and absence of subarticular tubercles (tubercles present, pads absent in A. baturitensis ) and large size, up to 23 mm SVL (largest A. baturitensis 16.3 mm).
Adelophryne patamona can be distinguished from A. maranguapensis by its posterior cloacal opening, without a transverse flap (flap present, cloaca opening ventrally in A. maranguapensis ) and large size, up to 23 mm SVL (largest A. maranguapensis 17.4 mm).
Adelophryne patamona can be distinguished from A. gutturosa , with which it occurs sympatrically, by the following characters: second phalange of Finger IV relatively long, 2X its width (short, length equal to or slightly greater than width in A. gutturosa ); canthus rostralis concave (straight); vomerine teeth 15–16 (2–8); large size, up to 23 mm SVL (largest A. gutturosa 16 mm) ( Fig. 1 View FIGURE 1 ). In the field these species can be easily distinguished by their distinctly different advertisement calls (see below). Characters useful for distinguishing among species of Adelophryne are in Table 1 View TABLE 1 .
Description of the holotype. A male, 17 mm SVL, head slightly longer than wide, as wide as adjacent part of body, head depth 50% of width. Snout rounded in dorsal and lateral profile. Distance between snout tip and eye slightly less than eye diameter, eye-nostril distance slightly less than internarial distance, 2X distance from nostril to snout tip, 75% of distance from eye to snout tip. Canthus rostralis indistinct, rounded, concave; loreal region sloping steeply to upper lip. Nostrils not projecting, round, directed laterally. Interorbital distance 1.3X internasal distance, 2X upper eyelid width. Pupil horizontal. Temporal region vertical; tympanum small, distinct, round, 35% of eye diameter, surrounded by a distinct annulus, partially obscured by a supratympanic fold, separated from eye by a distance equal to tympanum diameter. Low glandular ridge from tympanum to arm insertion barely visible.
Choanae small, laterally placed; vomerine processes transverse, posterior to choanae, each bearing 15 teeth. Tongue narrow, sides parallel, attached anteriorly. Vocal slits small, flanking tongue, vocal sac large, subgular.
Skin of dorsum, venter, and limbs smooth with scattered small tubercles; flanks and rear of thighs areolate. Cloacal opening above midpoint of thigh, directed posteriorly, dorsal flap absent.
Hand with large flat oval palmar and thenar tubercles. Underside of fingers with large flat subdigital pads (1–1–2–1); no pads under ultimate phalanges. Fingers flattened, unwebbed. Tips of fingers bluntly pointed, bearing discs not wider than the adjacent phalange, discs laterally grooved. Finger lengths I<IV<II<III; Finger IV relatively long, free part more than half that of Finger III. Phalangeal formula 2–2–3–3, second phalange of Finger IV twice as long as wide ( Fig. 1 View FIGURE 1 ). Terminal phalanges of Fingers I, II and III T-shaped, that of Finger IV blunt. Distal portions of fingers surrounded by a transparent rim of skin ( Fig 2 View FIGURE 2 ).
Tarsus smooth, no ridge or tubercle. A large distinct oval flat inner and a smaller round conical outer metatarsal tubercle. Subdigital pads (1–1–2–3–2) large, flat, located just distally of digital articulations; no supernumerary pads. Toes flattened, unwebbed; toe V more slender than the others. Tips of toes dilated into small, narrow discs, those of Toes II, III, IV slightly wider than adjacent phalange, ending in asymmetrically pointed tips; those of Toes I and V equal to width of adjacent phalange, with rounded or bluntly pointed tips. Discs with lateral grooves. Toe lengths I<II<V<III<IV. Phalangeal formula 2–2–3–4–3; terminal phalanges Tshaped. Distal portions of toes surrounded by a transparent rim of skin, although this is much less prominent on the toes than it is on the fingers ( Fig 2 View FIGURE 2 ).
Heel of adpressed hindlimb reaches a point between eye and tip of snout. Heels overlap slightly when hindlimbs are flexed at right angles to sagittal plane of body. Shank length 57% of SVL.
Colour of holotype in life. Ground colour of dorsum, flanks and dorsal surfaces of limbs medium brown. Central part of dorsum dark brown, bounded by a black middorsal “)(”-shaped mark and interorbital bar. Dorsum with a few other small, scattered black marks. Loreal and temporal regions dark brown. A black bar extending from tip of snout through eye to flank; another black bar in the inguinal region. Dorsal surfaces of limbs with black crossbands. Throat, chest and underside of arms dark grey with white or pale blue spots. Venter, underside of legs and rear of thighs reddish-brown with white or pale blue spots. A black anal spot. Palpebrum not reticulated, a black band along its upper rim. Iris copper with a red ring around pupil ( Fig 3 View FIGURE 3 ). In preservative the venter becomes brownish-grey with white spots.
Variation. The female paratype from Wokomung is the largest specimen collected, SVL 23 mm ( Fig 4 View FIGURE 4 ). Paratypes from Maringma ( Figs. 5–7 View FIGURE 5 View FIGURE 6 View FIGURE 7 ) are one male 18.4 mm SVL, five females 20.7–21.5 mm SVL, and two juveniles (11.8 and 11.9 mm SVL). Dorsal skin smooth to granular; ventral tubercles variable; vomerine processes each bearing 16–18 teeth in adults, 2–3 teeth in juveniles.
Colouration is variable, not sexually dimorphic ( Figs. 4–7 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 ). Black dorsal markings are variable; some specimens are darker with more pale blue spots and more conspicuous tubercles on dorsum. In the specimens from Maringma the throat, chest and underside of arms are black with white or pale blue spots and a few reddish-brown dots; the venter, underside of legs and rear of thighs are reddish-brown, sometimes with very few white or pale blue spots ( Figs. 6–7 View FIGURE 6 View FIGURE 7 ). Juveniles resemble adults, but the upper arm is yellowish-orange and the dorsum has more pale blue spots; ventrally they are black with numerous pale blue spots ( Fig. 5 View FIGURE 5 C).
Vocalisation ( Fig. 8 View FIGURE 8 ; Tables 2, 4 View TABLE 4 ). The following description is based on a sample of six advertisement calls from a single male, IRSNB 1978, recorded on the southeast slope of Mount Maringma (05º 12' 16" N, 060º 34' 39" W, 1376 m), on 24 November 2007 at 14h15, air temperature 19.8°C.
Call parameter N Mean Range
Call duration (s) 6 0.63 0.584–0.712 Inter-call interval (s) 5 3.85 3.365–4.255 Duration of first note (s) 6 0.038 0.035–0.042 Duration of second note (s) 6 0.056 0.055–0.057 Duration of third note (s) 6 0.429 0.400–0.497 Inter-note interval between the first and the second note (s) 6 0.075 0.072–0.081 Inter-note interval between the second and the third note (s) 6 0.052 0.047–0.059 Note repetition rate (notes/s) 12 9.07 8.66–9.26
Emphasized frequency (Hz) 18 3255 3251–3269 Temporal structure. The advertisement call of Adelophryne patamona consists of a group of three soft, unpulsed, whistle-like notes (two short, followed by one long) produced in rapid succession ( Fig. 8 View FIGURE 8 ). Although a feeble isolated single whistle-like note was sometimes heard, we never observed the specimen emitting that note, and thus we cannot confidently attribute it to A. patamona . However it is possible that, like in A. gutturosa (see below), a shorter call is emitted when a female is very close to a male. Males typically call antiphonally and there was no overlapping of the calls.
Spectral structure. Three harmonics are developed, with the fundamental frequency dominating (range: 3251–3269 Hz; mean: 3255 Hz). The distribution of sound energy decreases progressively through the higher harmonics. The frequency rises abruptly at the very beginning of the notes ( Fig. 8 View FIGURE 8 ). The amplitude of the third note rises sharply to a peak at the beginning of the note then drops off rapidly, rising again, slowly, to a peak near the end of the note. A smaller peak in amplitude occurs at the end of the note ( Fig. 8 View FIGURE 8 ). Vocalisation parameters of three Adelophryne species are compared in Table 4 View TABLE 4 .
Pulses 2 4–5 high amplitude pulses followed by up 0
to 7 low amplitude pulses
Emphasized harmonic Second First First Emphasized frequency (Hz) 3200–3700 3896–4979 3251–3269 Natural history. The specimens were collected at the end of the dry season, mostly at night, in leaf litter under medium-canopy forest ( Fig. 5 View FIGURE 5 D). One female (IRSNB 1975) and the two juveniles (IRSNB 1976–7) were collected among rootlets at the base of a tree. One male, IRSNB 1978, was calling in the afternoon under leaves in a hole between roots. Many males were heard calling during afternoons and nights but were very difficult to find in their hidden calling sites. No enlarged ova were found in the females.
The species was also heard on Maringma at 1414 m, and in high-canopy forest near the village of Wayalayeng (05º 14' 11" N, 060º 31' 04" W, 678 m), but none were collected at these locations.
Distribution. Known only from the Pakaraima Mountains in western Guyana. The species likely occurs in Roraima State, Brazil, only 2 km from the Maringma location. The collecting locality at 1411 m is the highest elevation reported for any Adelophryne .
Relationships. Adelophryne patamona appears to be most closely related to A. gutturosa , with which it is sympatric and sometimes syntopic (see below).
Colour pattern may not be useful for distinguishing among species of Adelophryne . A dark “)(” -shaped dorsal marking is present in several Adelophryne ( A. baturiensis , A. gutturosa and A. maranguapensis ; Hoogmoed et al. 1994). The ventral pattern of dark background with light spots is also common in the genus.

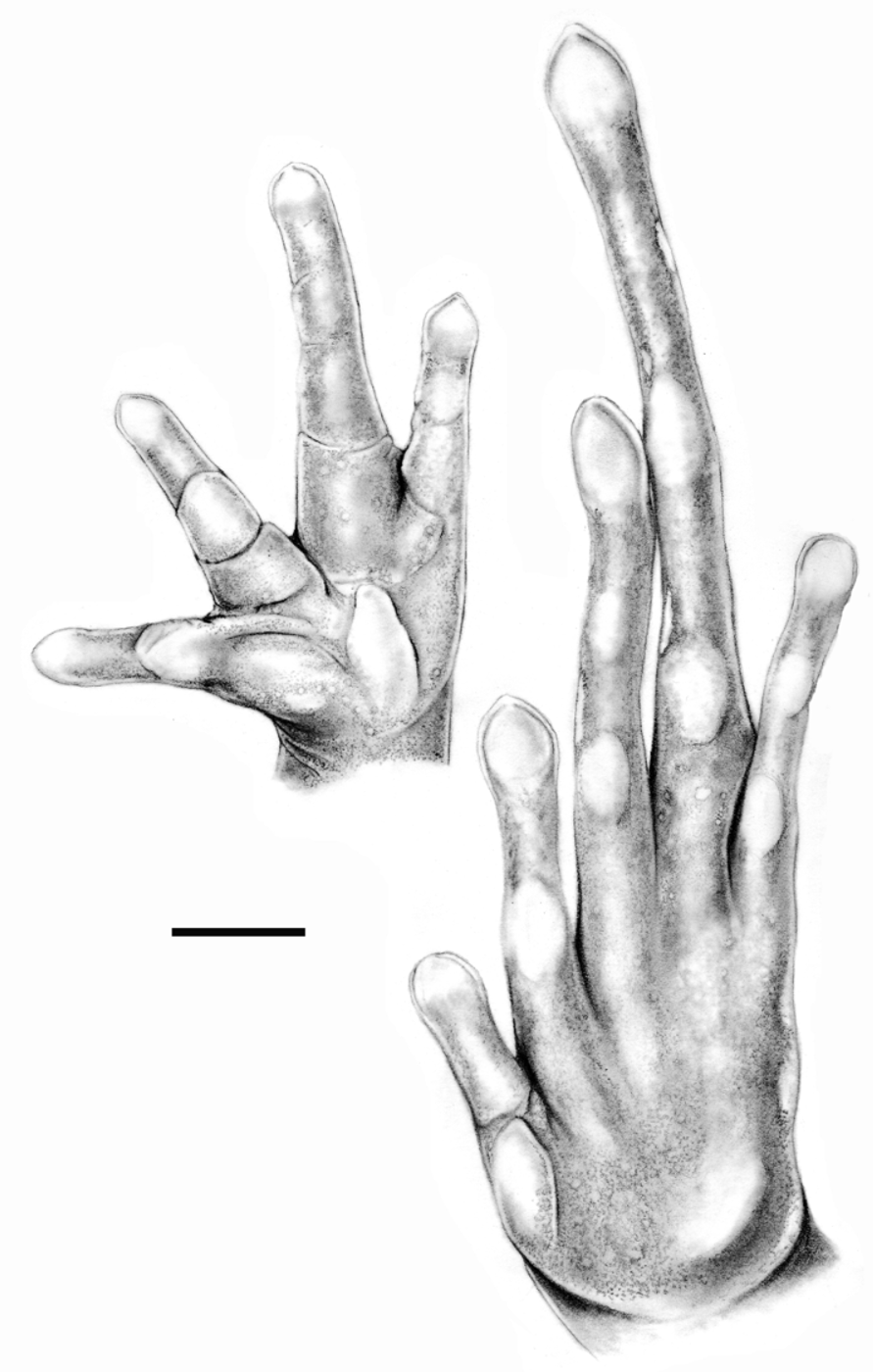






TABLE 1. Characters useful for distinguishing among species of Adelophryne. Some information comes from Hoogmoed and Lescure (1984), Hoogmoed et al. (1994), and Rodríguez and Duellman (1994).
| Species | Maximum SVL (mm) | Finger Pha- langeal Formula | Vo m e r i n e Teeth | Finger Pads or Tubercles | Dorsum | Distribution |
|---|---|---|---|---|---|---|
| adiastola | 14 | 2-2-3-2 | 2–8 | pads, 1-1-2-1 | granular | Amazonian Colum- bia and Perú |
| baturiensis | 16.3 | 2-2-3-3 | 3–8 | tubercles, 1-1-2-2 | smooth | Ceará, Brazil |
| gutturosa | 16 | 2-2-3-3 | 2–8 | pads, 1-1-2-1 | smooth/ shagreened | Guiana region |
| maranguapensis | 17.4 | 2-2-3-3 | 4–8 | pads, 1-1-2-2 | pustulous | Ceará, Brazil |
| pachydactyla | 11.1 | 2-2-3-2 | unknown | pads, 2-3-4-2 | smooth | Bahia, Brazil |
| patamona | 23 | 2-2-3-3 | 15–16 | pads, 1-1-2-1 | smooth/ shagreened, tubercles | Pakaraima Moun- tains, Guyana |
TABLE 4. Summary of some acoustic parameters of the advertisement calls of Adelophryne patamona sp. nov., A. gutturosa and A. adiastola. The advertisement call of A. adiastola was described by Heyer (1977, figs 3 - 4), under the name Phyzelaphryne miriamae (Hoogmoed & Lescure, 1984).
| Call parameter | A. adiastola | A. gutturosa | A. patamona sp. nov. |
|---|---|---|---|
| Number of notes | 1–10 | 2–15 | 3 |
| Note repetition rate (note/s) | 7 | 6.06–22.22 | 8.66–9.26 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.