
Limodromus hoffeinsorum, Schmidt, Joachim, 2015
|
publication ID |
https://doi.org/10.11646/zootaxa.3974.4.8 |
|
publication LSID |
lsid:zoobank.org:pub:62A22514-B276-4C50-8FE3-0CDABC7C62E3 |
|
DOI |
https://doi.org/10.5281/zenodo.6097838 |
|
persistent identifier |
https://treatment.plazi.org/id/03A4879C-1230-3D7C-FF34-CF6A329EFA39 |
|
treatment provided by |
Plazi |
|
scientific name |
Limodromus hoffeinsorum |
| status |
sp. nov. |
Limodromus hoffeinsorum View in CoL sp. n.
Figs. 1–11 View FIGURES 1 – 10 View FIGURE 11 .
Holotype. Male in Baltic amber; size of amber piece 14.0 x 12.0 x 4.8 mm, embedded in synthetic resin ( Hoffeins 2001), No. 1648- 3 in the collection of Christel and Hans Werner Hoffeins, Hamburg, Germany ( CCHH) (later to be deposited at the Senckenberg Deutsches Entomologisches Institut, Müncheberg, Germany) ( Fig. 1 View FIGURES 1 – 10 ).
Preservation status. Only the anterior part of the beetle body is conserved; the abdominal segments posterior of the hind coxae, the distal part of elytra, and the metatarsi are lost. The inclusion is surrounded by a reddishcoloured corona as a result of corrosion of the opened body cavity because the amber was in long-term contact to oxygen after sedimentation. Dorsal and ventral sides of head, pronotum and anterior half of hind body as well as antennae and legs (without metatarsi) are well visible in most of the morphological details of diagnostic importance.
Syninclusions. 1 expl. Chironomidae , 1 expl. Diptera (remains: legs, one wing), several plant trichomas or stellate hairs.
Description. Body length: Approx. 9.2 mm (head + pronotum = 3.8 mm).
Colour: Body blackish brown, shiny, apparently without metallic luster; palpi, antennae and legs not distinctly lightened.
Microsculpture: Discs of head and pronotum appearing polished with micro-meshes not visible even under magnification of 100 x (artifact?). Labrum with distinct isodiametric sculpticells ( 50 x). Surfaces of elytral intervals with very fine but distinct transverse meshes ( 50 x), which are regularly arranged ( Fig. 6 View FIGURES 1 – 10 ; elytra appearing finely fasciated under magnification below 50 x).
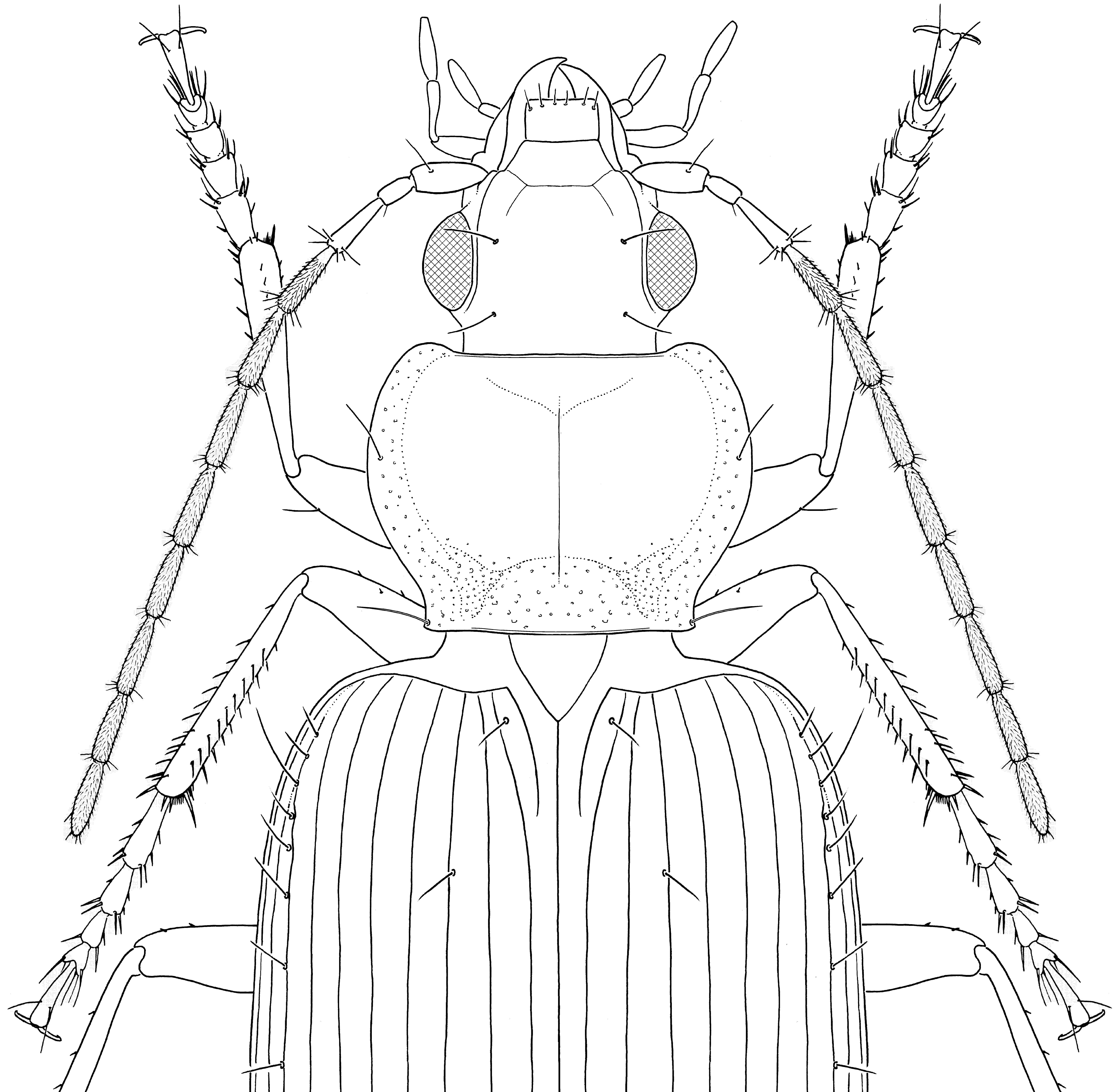
Head: Normal-sized, smooth; disc without reddish spots, convex, markedly constricted towards neck. Mandibles moderately slender, with external scrobe asetose. Eyes large and hemispherically protruded; temples relatively long, slightly more than 1/3 eye length, moderately wrinkled to the neck; supraorbital furrow moderately deep, extended to the posterior supraorbital seta; two supraorbital setae present. Mentum tooth simple, mentum setae small, inserted very close to each other immediately basad of mentum tooth, submentum with two pairs of lateral setae, outer pair very small ( Fig. 5 View FIGURES 1 – 10 ). Penultimate article of maxillary palpus bisetose in middle. Antennae slender, 1.45 times longer than head + pronotum; scape not enlarged, same length as third antennomere, with one seta on dorsal side; pedicel half as long as scape, with a single apical seta on ventral side; antennomere III without setae in addition to the normal ring of apical setae ( Fig. 4 View FIGURES 1 – 10 ).
Prothorax: Pronotum 1.32 broader than head, smooth, disk convex, in dorsal view cordate, transverse (width/ length = 1.20), broadest somewhat anterior of middle, with basal margin 1.13 times broader than apical margin. Sides evenly rounded in anterior 2/3 and concave before base. Anterior margin finely beaded throughout, feebly concave, with front angles slightly protruded, widely rounded. Posterior margin finely beaded throughout, almost straight, with hind angles very slightly shifted anteriad; latter slightly acute, slightly protruded laterally. Median longitudinal impression moderately deep in middle, disappearing near apex and base, both anterior and posterior transverse impressions distinct. Lateral gutter wide throughout, near apex almost as broad as antennal scape, slightly widened towards base, laterobasal depressions very broad and deep; rather coarsely and densely punctuate, lateral gutter more sparsely punctuate. Both lateral and laterobasal setae present, with laterobasal setae located on margin. Proepisternum smooth, prosternal process not marginated; anterior coxal cavities closed posteriorly.
Pterothorax (anterior portion): Elytra moderately convex, presumably sub-parallel in dorsal view, distinctly wider than head. Shoulders broad, humeral angles rounded, basal groove moderately concave, protruded towards scutellum and humerus as well. All striae deeply impressed, impunctate, parascutellar stria developed, intervals moderately convex. Parascutellar seta and anterior seta on third interval present, latter adjoined to the third stria; umbilicate series of the anterior elytral half consists of 10 setae adjoining eighth stria of which the first seven setae are moderately separated within the humeral group; third seta more than two times longer than second and fourth setae. Metepisternum with outer margin almost two times longer than anterior margin, smooth ( Fig. 3 View FIGURES 1 – 10 ). Hind wings fully developed.
Legs: Mesocoxa with a single ridge seta; metacoxa bisetose, with inner seta absent ( Fig. 7 View FIGURES 1 – 10 ). All trochanters with seta. Pro-, meso- and metafemur each with two ventral setae; metafemur with several short subapical setae. Protibia not markedly broadened towards apex, with antennal cleaner organ situated subapically, with almost complete groove on dorsal surface ( Fig. 8 View FIGURES 1 – 10 ), and with inner apical spur 2/3 as long as first protarsomere ( Fig. 10 View FIGURES 1 – 10 ). Dorsal surface of pro- and mesotarsomeres glabrous beside normal setation, without internal and external sulci. Proximal three protarsomeres moderately broadened (male!), each with two complete, dense rows of broad adhesive hairs ( Fig. 9 View FIGURES 1 – 10 ). Ventral lobes of fourth protarsomeres short, almost symmetric ( Figs. 9, 10 View FIGURES 1 – 10 ). Dorsoapical setae present on both sides of tarsomeres I–II, lacking on III and IV; lateroapical setae on both sides of tarsomeres III and IV present ( Fig. 10 View FIGURES 1 – 10 ). Tarsomeres V glabrous on ventral side.
Etymology. The new species is dedicated to Christel and Hans Werner Hoffeins, Hamburg, for their diverse contributions to the study of arthropods in amber.
Species group recognition and differential diagnosis. Tribus: A species of the tribe Platynini based on the combination of the following characters: 1) mandibular scrobe asetose; 2) penultimate article of maxillary palpus bisetose; 3) two pairs of supraorbital setae present in normal position; 4) head and pronotum without stridulating mechanism; 5) elytra with dorsal setae in third interval; 6) elytra with apical margin entire, not truncate; 7) elytra without internal plica; 8) prosternal process not marginated; 9) anterior coxal cavities closed posteriorly; 10) mesepimeron not reaching the mesocoxal cavities (= mesocoxal cavities conjunct); 11) metepimeron lobe-like dilated; 12) protibia not markedly broadened towards apex, with 13) antennal cleaner organ situated subapically and 14) with groove almost complete on dorsal surface; 15) claws simple, not pectinate.
Remarks: The systematic position of the Platynini within the “modern Carabidae ” (= Harpalinae sensu auct.) is still largely unsettled. They are often considered the sister group of Sphodrini (e.g., Habu 1978, Casale 1988, Lorenz 2005, Bousquet 2010), or even the sister group of a single branch of the latter tribe (Synuchi: Lindroth 1956), however, not a single synapomorphy is known that supports these assumptions. In addition, although representatives of the Platynini tribe are relatively easy to recognize (e.g., based on the combination of the character states mentioned above) the currently known diagnostic characters of Platynini seem to be plesiomorphic as well. The combination of the above listed characters 4, 8, 14, and 15, is suitable for differentiation of the fossil specimen from species groups of Sphodrini .
Genus: The new fossil species is a representative of the genus Limodromus based on the combination of the following characters: 1) dorsal and ventral surfaces of body glabrous beside normal setation; 2–4) head capsule completely blackish without reddish brown spots, without punctures on disc, and with a distinct constriction towards neck; 5) eyes large and hemispherical; 6) mandible normal, not markedly slender; 7) mentum tooth developed, simple; 8) mentum setae small, inserted very close to each other immediately basad of mentum tooth; 9) third antennomere glabrous, only with a ring of primary setae apically; 10–13) pronotum distinctly broader than head, cordate with laterobasal angles large, with lateral gutter and laterobasal grooves very broad, and with laterobasal seta present; 14) elytra with anterior dorsal seta in third interval present; 15) metacoxa bisetose (inner seta absent); 16) ventral lobes of fourth protarsomeres short, almost symmetric.
Remarks: Monophyly of Limodromus is well supported based on at least four synapomorphies of which three are features of the male genital armature ( Schmidt 2000, 2005). This cannot be determined in L. hoffeinsorum sp. n. because the abdomen is lost in the fossil specimen. The position of the mentum setae as observed in L. hoffeinsorum sp. n. and in other Limodromus species is the fourth proposed synapomorphic character but it is also present in the following four lineages of Platynini :
Clade of Agonum Bonelli, 1810 + Agonidium Jeannel, 1948 (see Liebherr & Schmidt 2004): In general, species of this clade are easily to recognize based on the derived shape of pronotum with +/- rounded hind angles. In addition, species of Agonidium differ by the antennal pedicellus bearing a ring of setae at apex, species of Europhilus Chaudoir, 1859 differ additionally by the pubescent distal portion of third antennomere, and both the latter taxa as well as Agonum s. str. differ by the very narrow anterior half of the pronotal marginal gutter. In all other lineages of Agonum s.l. (summarized in the subgenus Agonothorax Motschulsky, 1850 = Olisares Motschulsky, 1865 , see Liebherr & Schmidt, 2004, Liebherr et al. 2005), the mentum setae are inserted laterad of the base of the mentum tooth, and thus they are different from Limodromus .
Anchomenus View in CoL clade sensu Liebherr (1991): Within this clade only the western Palearctic species of Anchomenus View in CoL ( A. cyaneus Dejean, 1828 View in CoL , A. dorsalis Pontoppidan, 1763 View in CoL , A. kurnakovi Kryzhanoviskij, 1983 View in CoL , and the A. dohrnii View in CoL species group sensu Schmidt 2014), as well as some species of Elliptoleus Bates, 1882 View in CoL , and Sericoda Kirby, 1837 View in CoL have the mentum setae similarly developed as in Limodromus spp. However, Anchomenus View in CoL species differ by the antennal pedicellus bearing a ring of setae at apex. Species of Elliptoleus View in CoL and Sericoda View in CoL are easily to recognize by the much smaller body size and by the shape of the pronotum with fully rounded laterobasal angles.
Paranchodemus Habu, 1978 View in CoL : Species of this genus differ by the densely punctuate neck, bifid mentum tooth, and very narrow pronotal lateral gutter.
Platynus Bonelli, 1810 View in CoL : Externally, species of this genus differ in having a pair of reddish brown spots on neck.
The presumably plesiomorphic character state (mentum setae located distinctly laterally of the base of the mentum tooth) is observed in all the remaining species groups of Platynini View in CoL including the highly diverse lineages related to the Pantropic Colpodes Macleay, 1825 View in CoL ( Colpodes View in CoL sensu lato in sense of Schmidt 2000), the Afro-Oriental Metagonum Jeannel, 1948 View in CoL , the enigmatic and probably ancient genera Atranus LeConte, 1848 View in CoL , Meleagros Kirschenhofer, 1999 View in CoL , and Tarsagonum Darlington, 1952 View in CoL , and thus in the majority of platynine beetles.
Differentiation from modern Limodromus species: The Eocene L. hoffeinsorum sp. n. differs from the northern Holarctic species of this genus [= assimilis species group (Schmidt 2005): L. assimilis (Paykull, 1790) , L. decentis (Say, 1823) , L. indecentis ( Liebherr & Will, 1996) , L. krynickii (Sperk, 1835) , L. longiventris (Mannerheim, 1825) , L. subovatus (Putzeys, 1875] by smaller body size, regular transverse pattern of the elytral microsculpture, two pairs of setae on the submentum, more markedly concave pronotal margin near base, and a larger number of adhesive hairs on male protarsomeres I–III. In external shape, body size, and microsculpture, the new species is more similar to L. klausnitzeri Schmidt, 2005 from northern Vietnam, however, it is easily to be distinguished by the longer temples, presence of a single seta on the pedicellus, pronotal posterior margin finely beaded throughout, metacoxae bisetose, dorsal surfaces of all tarsomeres without longitudinal furrows, third tarsomeres without dorsoapical setae, and fifth tarsomeres glabrous on ventral surface.
Relationships. With respect to the modern species, L. klausnitzeri was hypothesized the most basal representative of the genus Limodromus , and eight synapomorphies have been identified which define the monophyly of the remaining six species from the northern Holarctic (= assimilis species group, Schmidt 2005). The Eocene L. hoffeinsorum sp. n. seems to occupy an intermediate position between both lineages based on the following arguments:
i) As in L. klausnitzeri , the new species has two pairs of setae on submentum. This character state is doubtless a symplesiomorphy in Platynini View in CoL ; reduction of the outer pair of setae in the stem lineage of the assimilis group is highly probable.
ii) L. hoffeinsorum sp. n. possesses at least five derived characters together with the assimilis group (Schmidt 2005): 1) temples relatively long; 2) antennal pedicellus with a single seta; 3) metacoxae bisetose (inner seta reduced); 4) longitudinal furrows on tarsal surfaces reduced; 5) dorsoapical setae on fourth tarsomeres reduced.
iii) L. hoffeinsorum sp. n. and L. klausnitzeri both are characterized by finely and regularly transverse microsculpture patterns on elytra, while the other species possess isodiametric sculpticells. Transverse meshes represent the derived character state of the group ( Liebherr & Schmidt 2004, Schmidt 2005).
Because L. hoffeinsorum sp. n. shares the majority of derived character states with the northern Holarctic species, it seems most likely that the new species is a representative of this lineage. Here it represents the most basal branch (= sister taxon of the assimilis group) characterized by two pairs of setae on submentum while the outer pair was reduced in the evolution of the modern species. Based on this hypothesis parallel evolution of transverse patterns of the elytral microsculpture has to be assumed for L. hoffeinsorum sp. n. and L. klausnitzeri .
These conclusions are provisional and have to be confirmed based on additional material, particularly by the investigation of male and female genital characters of the fossil species. The latter is possible since non-invasive 3- dimensional computerised X-ray microscopy allows examination of even minute internal structures of arthropods (e.g., Dierick et al. 2007, Greco et al. 2008, Kehlmaier et al. 2014). It would be therefore very exciting if additional Limodromus fossils were detected in the large collections of Baltic amber in future.
Biogeographical considerations. The occurrence of Limodromus in Baltic amber is in accordance with the modern distribution of the genus. The suspected distributional area of the Baltic amber producing forests was situated in the Eocene paratropical floral region of the northern hemisphere (see Mai 1995, Weitschat & Wichard 1998). The flora of this region had a close relationships to the modern flora of Indo-Malaya ( Wolfe 1975). In this respect it is worthy to note that a representative of the presumably most ancient branch of the genus Limodromus was found to occur in the mountains of northern Vietnam (Schmidt 2005). The modern climate of this area is characterized as subtropical to warm-temperate and hence, corresponding to the paratropical climate in sense of Wolfe (1975, see Graham 2011). It can thus be assumed that the habitat of L. hoffeinsorum sp. n. was similar to that of the modern L. klausnitzeri . The sister group of L. hoffeinsorum sp. n. is distributed from the warm-temperate to the boreal zone across the Holarctic region. This lineage ( assimilis group) has adapted to somewhat colder climates probably in the course of the climate cooling after the Eocene optimum. The Holarctic distribution of the modern Limodromus becomes apparent in light of the fact that the temperate floras of North America, western Eurasia and East Asia were almost continuously connected until the Miocene ( Mai 1995). Up to this age the Holarctic might have been a continuous faunal region, too, at least for winged species with high dispersal power.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Tribe |
Platynini |
|
Genus |
Limodromus hoffeinsorum
| Schmidt, Joachim 2015 |
L. klausnitzeri
| Schmidt 2005 |
Meleagros
| Kirschenhofer 1999 |
L. indecentis (
| Liebherr & Will 1996 |
A. kurnakovi
| Kryzhanoviskij 1983 |
Paranchodemus
| Habu 1978 |
Tarsagonum
| Darlington 1952 |
Metagonum
| Jeannel 1948 |
Elliptoleus
| Bates 1882 |
Atranus
| LeConte 1848 |
Sericoda
| Kirby 1837 |
L. krynickii
| Sperk 1835 |
A. cyaneus
| Dejean 1828 |
Colpodes
| Macleay 1825 |
L. longiventris
| Mannerheim 1825 |
L. decentis
| Say 1823 |
Platynus
| Bonelli 1810 |
L. assimilis
| Paykull 1790 |
A. dorsalis
| Pontoppidan 1763 |