
Sperosoma obscurum Agassiz and Clark, 1907
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4092.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:EA66CAE5-F6CE-44BA-A5FF-67F2BEE6DEE8 |
|
DOI |
https://doi.org/10.5281/zenodo.6055438 |
|
persistent identifier |
https://treatment.plazi.org/id/03A4AB67-FFE0-FF87-FF1D-FC34FD836029 |
|
treatment provided by |
Plazi |
|
scientific name |
Sperosoma obscurum Agassiz and Clark, 1907 |
| status |
|
Sperosoma obscurum Agassiz and Clark, 1907 View in CoL
( Figures 1–5 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 )
Material examined. 69 specimens from 50 records (NIWA, Te Papa, MV).
Chatham Rise/East Coast South Island: 3 specimens (frozen), 44° 42.0´S, 173° 15.9´E, 911–948 m, NIWA78364; 4 specimens, 44° 42.0´S, 173° 15.9´E, 911–948 m, NIWA78364; 2 specimens (dry, 200, 200 mm TD), 44° 13.8´S, 178° 26.4´E, 1090 m, NIWA23712; 2 specimens (dry, 116, 145 mm TD), 43° 37.5´S, 174° 16.4´E, 896– 890 m, NIWA23714; 1 specimen (dry, 150 mm TD), 42° 42.9´S, 179° 57.5´W, 950–1070 m, NIWA25960; 1 specimen (210 mm TD), 44° 41.0´S, 174° 20.9´E, 859 m, NIWA29426; 1 specimen (210 mm TD), 44° 41´S, 174° 20.9´E, 859 m, NIWA29440; 1 specimen (183 mm TD), 42° 47.8´S, 179° 57.1´E, 985 m, NIWA29480; 1 specimen (220 mm TD), 44° 41.1´S, 176° 37.1´W, 863 m, NIWA29481; 1 specimen (205 mm TD), 44° 38.0´S, 176° 58.6´W, 848–898 m, NIWA29482; 1 specimen (220 mm TD), 44° 37.4´S, 176° 4.6´W, 800 m, NIWA29488; 1 specimen (dry, 190 mm TD), 44° 25.8´S, 178° 31.3´W, 868 m, NIWA29490; 1 specimen (225 mm TD), 44° 23.1´S, 178° 9.4´W, 805 m, NIWA29501; 1 specimen (200 mm TD), 44° 35.3´S, 177° 27.4´W, 852 m, NIWA29510; 1 specimen (196 mm TD), 42° 43.5´S, 178° 20.9´W, 900–924 m, NIWA33593; 1 specimen (182 mm TD), 42° 48.2´S, 179° 57.5´W, 958–1013 m, NIWA45104; 2 specimens (192 [dry], 210 mm TD), 42° 50.2´S, 179° 58.6´W, 908– 904 m, NIWA58237; 2 specimens (1 dry; 192, 205 mm TD), 42° 46.4´S, 179° 27.6´W, 953– 942 m, NIWA58238; 1 specimen (195 mm TD), 42° 46.4´S, 179° 27.6´W, 953– 942 m, NIWA58244; 1 specimen (200 mm TD), 42° 46.4´S, 179° 27.6´W, 953– 942 m, NIWA58245; 1 specimen (145 mm TD), 44° 1.7´S, 174° 35.5´W, 796–882 m, NIWA58622; 1 specimen, 43° 18.0´S, 174° 12.0´W, 995– 967 m, Te Papa EC9648. West Coast North Island: 1 specimen (171 mm TD), 36° 6.0´S, 173° 16.0´E, 850– 850 m, NIWA29509; 1 specimen, 37° 28.5´S, 173° 58.6´E, Te Papa EC3978. East Coast North Island: 1 specimen (133 mm TD), 39° 43.6´S, 178° 13.2´E, 957 m, NIWA19345; 1 specimen (158 mm TD), 39° 50.0 S, 177° 39.0´E, 1000–1100 m, NIWA29513; 1 specimen (100 mm TD), 39° 27.5 S, 178° 25.3´E, 796– 796 m, NIWA45083; 2 specimens (193, 205 mm TD), 39° 38.0 S, 178° 1.3´E, 862 m, NIWA76745. Challenger Plateau: 1 specimen (190 mm TD), 37° 30.8´S, 168° 35.4´E, 966 m, NIWA29522; 1 specimen (dry, 120 mm TD), 39° 56.3´S, 168° 16.8´E, 832–834 m, NIWA34841; 1 specimen (dry, 142 mm TD), 39° 45.2´S, 168° 2.1´E, 831–840 m, NIWA34842; 3 specimens (160, 165, 165 mm TD), 41° 0.6´S, 169° 6.0´E, 914 m, NIWA45107; 1 specimen, 38° 56.6´S, 171° 3.0´E, Te Papa EC7502. Campbell Plateau: 1 specimen (dry, 174 mm TD), 48° 40.0´S, 170° 48.5´E, 814 m, NIWA29435; 2 specimens (207, 212 mm TD), 50° 24.4´S, 173° 44.0´E, 755 m, NIWA29438; 1 specimen (215 mm TD), 53° 41.1´S, 167° 47.3´E, 949– 947 m, NIWA46088; 1 specimen, 50° 15.1´S, 174° 5.5´E, 772–776 m, NIWA66703; 1 specimen (210 mm TD), 52° 30.8´S, 173° 22.7´E, 666–684 m, NIWA66707; 1 specimen (200 mm TD), 47° 27.2´S, 169° 58.9´E, 941–951 m, NIWA66712; 3 specimens, 47° 27.2´S, 169° 58.9´E, 758– 725 m, NIWA66714; 1 specimen, 48° 44.5´S, 172° 0.0´E, Te Papa EC3692. Lord Howe Rise: 1 specimen (225 mm TD), 33° 4.1´S, 162° 47.0´E, 809 m, NIWA45026; 1 specimen (104 mm TD), 32° 59.4´S, 163° 8.7´E, 830 m, NIWA45027; 1 specimen (165 mm TD), 35° 59.0´S, 166° 11.3´E, 723 m, NIWA45091. Puysegur Banks: 1 specimen (dry, 185 mm TD), 46° 37.3´S, 166° 34.7´E, 827 m, NIWA23713. Kermadec Ridge: 1 specimen (185 mm TD), 30° 36.5´S, 178° 22.5´W, 1030 m, NIWA46077. West Norfolk Ridge: 1 specimen, 35° 8.1´S, 169° 28.4´E, 868–872 m, MV (unregistered); 3 specimens, 33° 37.1´S, 166° 55.5´E, 1017–1042 m, MV (unregistered); 2 specimens, 33° 37.1´S, 166° 55.5´E, 1017–1042 m, MV F167728. Southern Australia: 1 specimen, 34° 40.2´S, 120° 24.7´E, 1110– 1130 m, MV (unregistered); 2 specimens, 44° 5.9´S, 147° 24.6´E, 810–1020 m, AM J.24886. Unknown location: 2 specimens, Te Papa EC6900. Unless stated, all stored in 80% ethanol.
Size range. The median test diameter of the 43 specimens measured was 190 mm, with a total size range of 29–225 mm TD. This is very similar to the size range of the 39 specimens originally collected by the Albatross in 1902 (20–220 mm TD).
Occurrence. The type material and other initial specimens of S. obscurum were collected from off the islands of Hawaii in 302–1266 m. There appear to have been no subsequent captures reported, although submersible observations were recorded from south of Maui Island in 1475–1935 m (Malahoff et al. 1988). The material examined here extends the species range to New Zealand and South Australia. The species is widespread around New Zealand, with numerous records from the Chatham Rise, the east coasts of North and South Islands including the Bay of Plenty, the Challenger Plateau, the Lord Howe Rise, and Kermadec and West Norfolk Ridges. The two records from Australia are from the southwest (off Albany) and southeast (Tasmania) ( Figure 2 View FIGURE 2 ).
The conservative depth range for the New Zealand and Australian specimens (based on the shallowest maximum depths and deepest minimum depths from records for which two values were recorded) is 684–1017 m, with a maximum potential range of 375–2130 m.
Remarks. The specimens examined agree well with the descriptions of Agassiz and Clark (1907) and Mortensen (1935), in particular with regard to the most distinguishing feature of the species—the tridentate pedicellariae ( Figure 3 View FIGURE 3 ). There is only a single form of this type, although they come in a range of sizes (approximately 0.5–2.5 mm) and their long narrow blades readily distinguish them from other species of Sperosoma . The triphyllous pedicellariae, although in general being less variable between species than the tridentate type, also show close agreement—the blades about 0.5 mm long with variously truncated and rounded tips, the basal involution of the blade not well developed. Ophicephalous pedicellariae were not seen, these forms thus far having only been found in S. durum .
Coronal plate counts were made on four specimens, and compared with those of the type specimens examined by Agassiz & Clark (1907) and Mortensen (1935) ( Table 2 View TABLE 2 ). The 15–17 ambulacral plates and 10–11 interambulacral plates on the oral side, and the 21–22 ambulacral plates on the aboral side match perfectly with the types, the only difference being an additional 2–3 plates in the aboral interambulacra of the new material. However, aboral plate boundaries were difficult to trace so these counts may be less reliable in any case. The ambulacra are slightly wider than the interambulacra at the ambitus (61 mm vs 58 mm in NIWA58244) as expected in Sperosomatinae .
The unusual arrangement of the oral ambulacra, unique to Sperosoma , in which the main plate is divided into an outer plate and an inner, non-poriferous plate separated by the secondary plates (Koehler 1897) is clearly seen in these specimens ( Figure 4 View FIGURE 4 ). The ambulacral pores are thus limited to the outer main plate and the secondary plates, on which they are found near the distal margin. Examination of the inner surface of these non-poriferous plates in NIWA58245 showed no sign of any lateral canals leading towards them or any evidence on them of undeveloped pores or podia.
The plate arrangement in the aboral ambulacra is quite different in that the main plate is generally not divided and spans most or all of the entire column ( Figure 4 View FIGURE 4 ). The elongated outer secondary plate frequently reaches the adradial margin and the inner plate is enclosed by the other two plates.
The transition of the plate arrangement between the aboral and oral surfaces occurs abruptly at the ambitus where consecutive inner secondary plates start to come in contact with each other, thus dividing the main plate. The outer portion of the main plates subsequently widen and expand to completely occupy the outer third of the column, while the outer secondary plates decrease in size and become restricted to the centre of the column, in broad contact with the inner secondary plates ( Figure 5 View FIGURE 5 ). This observation demonstrates that the assertion by Mooi et al. (2004) that Mortensen (1935) and Koehler (1897) had misinterpreted “cracks” in the soft test of Sperosoma grimaldi and other species as sutures separating the main ambulacral plate into poriferous and non-poriferous components is only partially correct, as it appears to have been based on examination of the aboral side only. While Mortensen at least partly misinterpreted the plate structure in the aboral ambulacra (as explained by Mooi et al. 2004, and compare Figure 4 View FIGURE 4 here with Mortensen 1935, Fig 107), there is no doubt about the existence of nonporiferous plates in the oral ambulacra, and the apparent breach of the Ocular Plate Rule (Mooi and David 1998, Mooi et al. 2004) in this genus.
The oral surface of the specimens examined are consistently covered with small tubercles of a similar size, especially around the peristome (as described by Agassiz and Clark 1907), whereas there is some variation in the arrangement of the primary tubercles: Agassiz and Clark (1907) state that there is a primary tubercle on each ambulacral plate near the ambitus whereas Mortensen’s (1935) specimen has them only on every other plate. In NIWA58244 there is a primary tubercle on each of these plates, but in NIWA58245 the occasional plate is missing a tubercle and in NIWA33593 the arrangement is more like every second plate as in Mortensen’s specimen. The tubercle arrangement in the oral interambulacra is more constant, with an outer series decreasing in size most of the way to the peristome and a shorter inner series restricted to the plates nearest the ambitus.
Some colour variability between specimens was noted, from the usual intense purple through to lighter shades of purple and also tan, as seen in a specimen examined from the Kermadec Ridge (NIWA46077). Colour variability in this species was noted by Agassiz & Clark (1907), who observed gray or yellowish-brown specimens among those of a more typical purple shade. Test plates of the cleaned specimen were outlined fairly distinctly by white connective tissue—although Agassiz and Clark (1907) describe this tissue as being darker than the test in the Hawaiian specimens their figures seem to show otherwise (as pointed out by Mortensen 1935). The explanation for this variability may be that the white connective tissue only becomes apparent when the delicate layer of skin covering the plates and sutures is removed, either during capture and preservation or by manual or chemical (bleach) removal.
Although very fragile now, the Kermadec specimen was unusually intact when collected, retaining most appendages including several oral spines with hoofs and others with a swelling in the shaft indicating the presence of a parasitic copepod ( Figure 4 View FIGURE 4 ). These copepods, in the genus Calvocheres , are also known from the spines of Sperosoma quincunciale , Calveriosoma gracile , and Hygrosoma hoplacantha (Mortensen 1903, Agassiz and Clark 1909, Stock 1968, Anton et al. 2013).

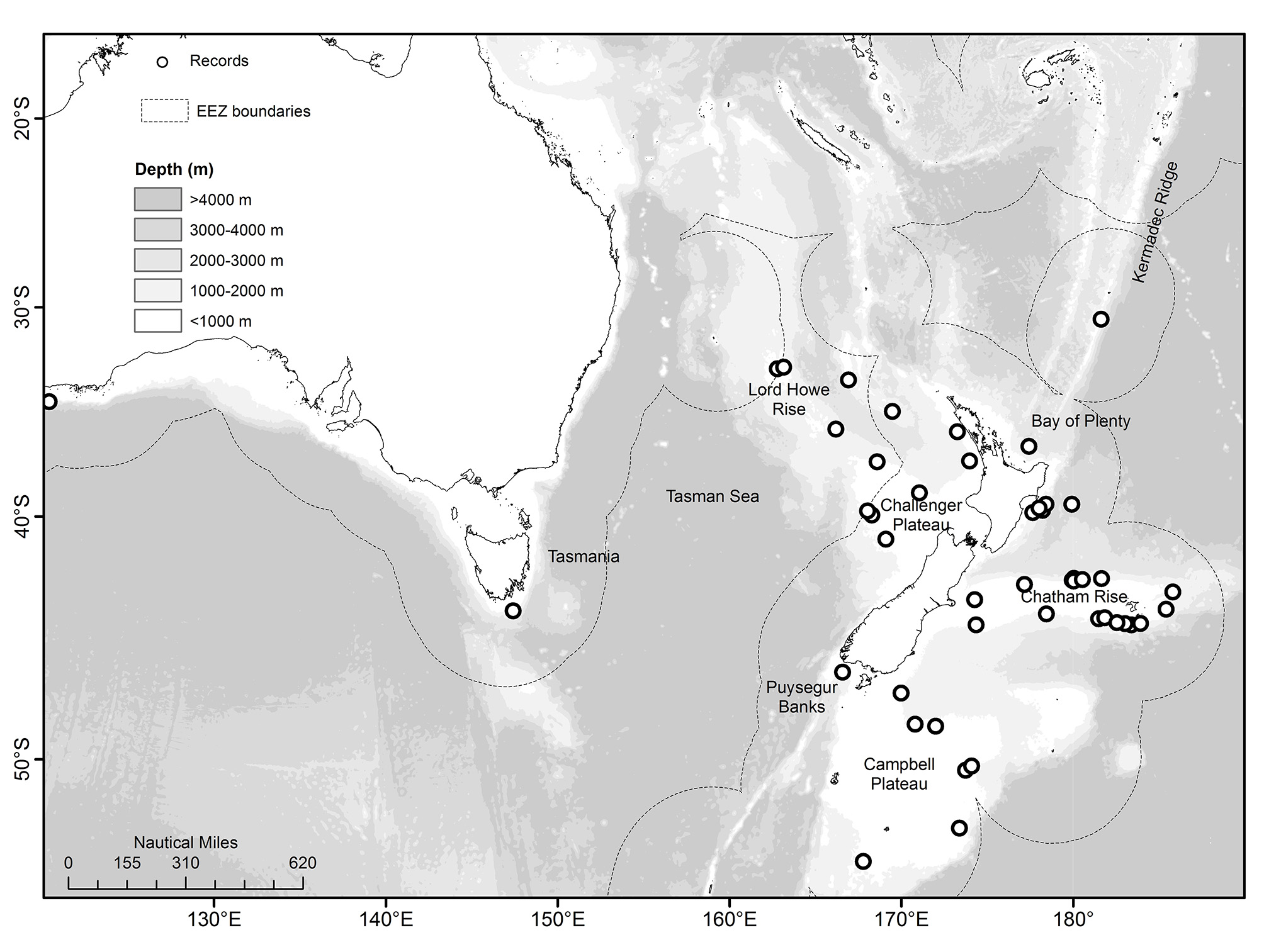
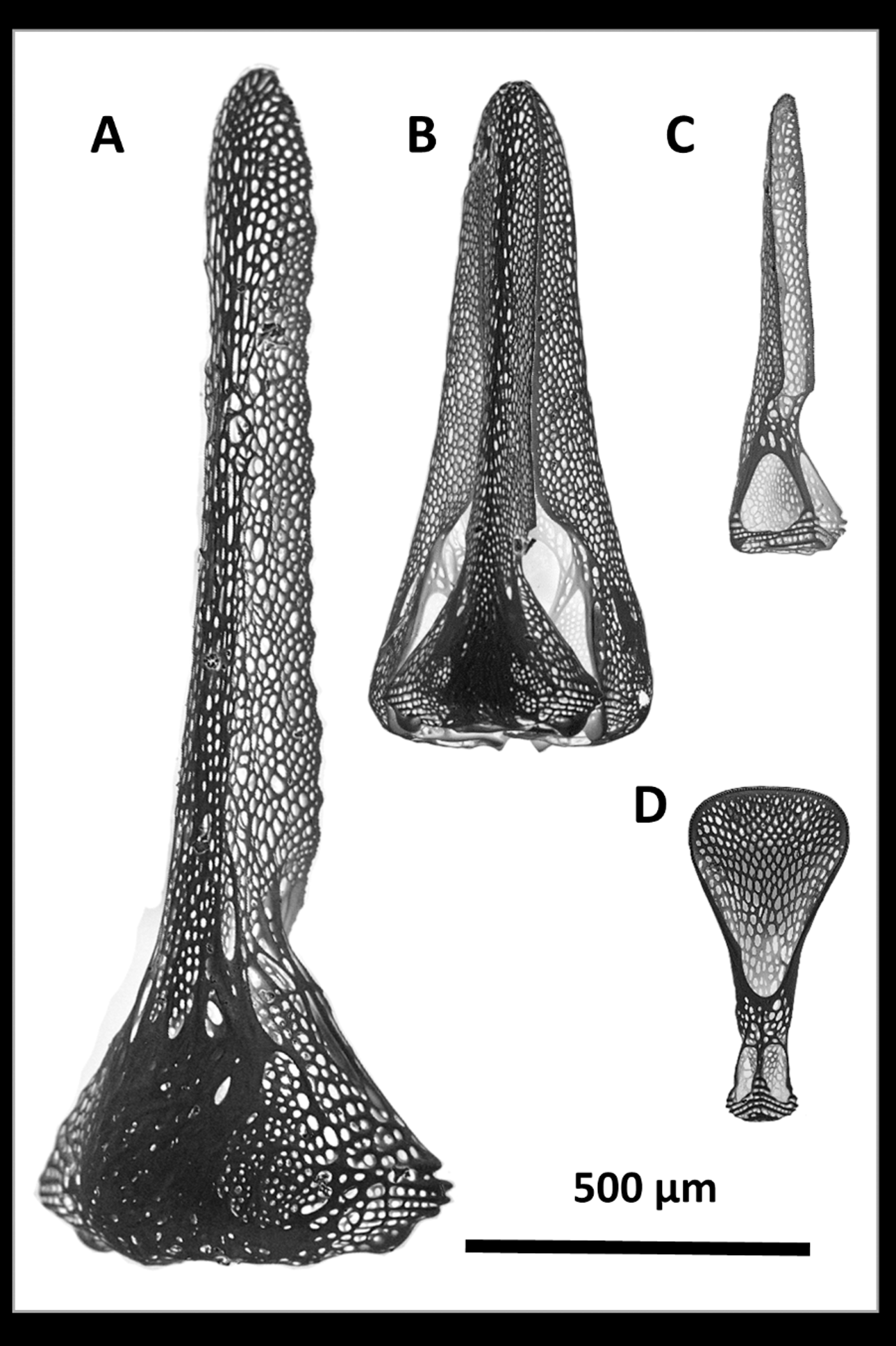
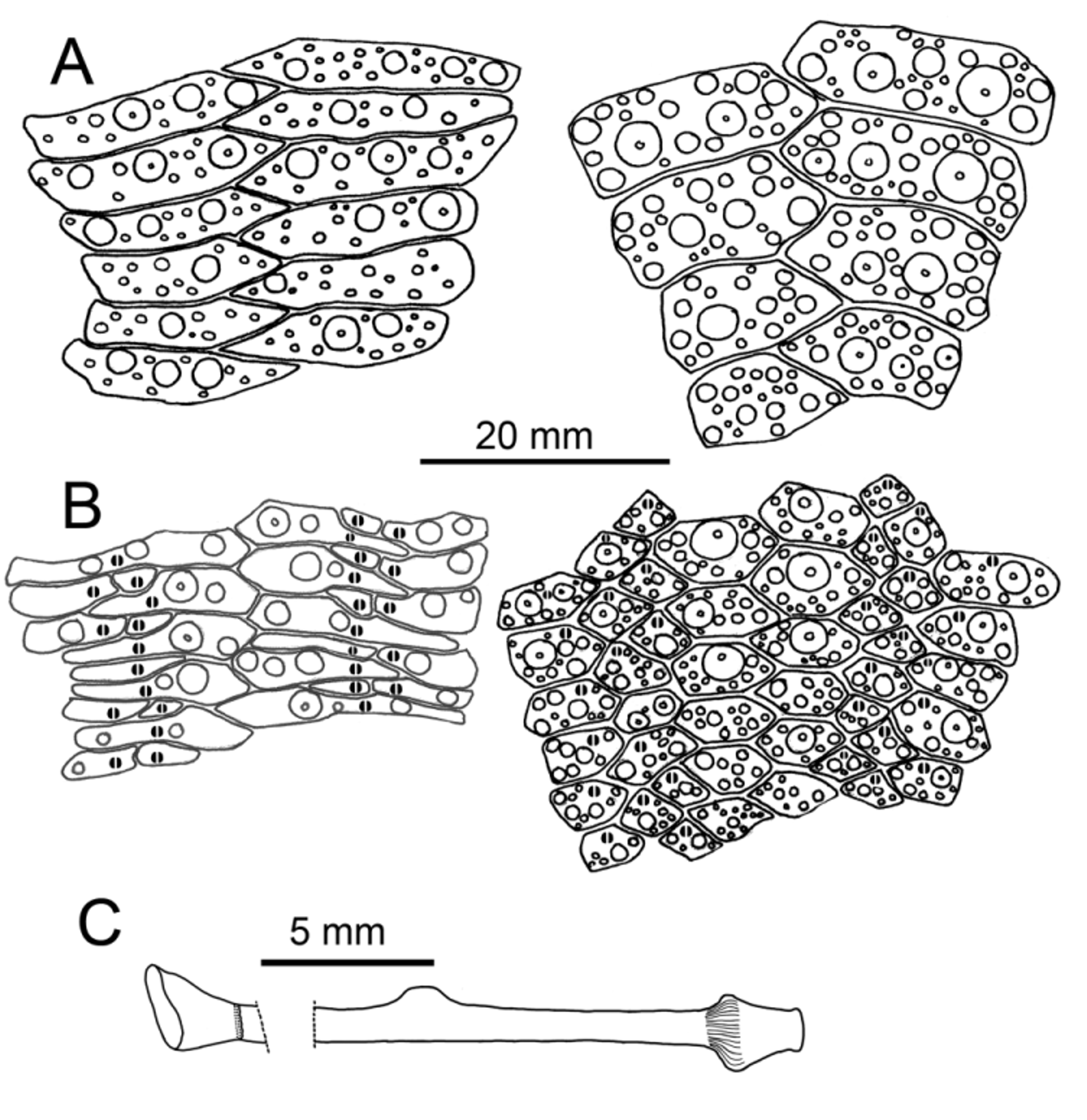

TABLE 2. Coronal plate counts for Sperosoma obscurum. –, not counted.
| Number of plates | ||
|---|---|---|
| oral aboral | ||
| Specimen | TD (mm) | A IA A IA |
| NIWA58244 | 195 | 17 10 22 22 |
| NIWA58245 | 200 | 15 11 21 21 |
| NIWA33593 | 196 | 16 11 – – |
| NIWA46077 | 185 | 16 11 – – |
| Agassiz and Clark (1907) | 169 | 18 13 25 19 |
| Mortensen (1935) | 130 | 16–17 10– 11 19–20 16–17 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.