
Hyloxalus arliensis, Acosta-Galvis & Vargas-Ramírez & Anganoy-Criollo & Ibarra & Gonzáles, 2020
|
publication ID |
https://doi.org/10.11646/zootaxa.4758.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:80FC5819-A32A-4223-9D6A-927578AEF910 |
|
DOI |
https://doi.org/10.5281/zenodo.3812885 |
|
persistent identifier |
https://treatment.plazi.org/id/03A4D47A-B740-6A46-FF15-DF124C6A51E7 |
|
treatment provided by |
Plazi |
|
scientific name |
Hyloxalus arliensis |
| status |
sp. nov. |
Hyloxalus arliensis sp. nov.
( Figures 4–6 View FIGURE 4 View FIGURE 5 View FIGURE 6 )
Hyloxalus “ Ibagué” Grant, Frost, Caldwell, Gagliardo, Haddad, Kok, Means, Noonan, Schargel & Wheeler, 2006. Holotype. An adult female ( Figure 4 View FIGURE 4 ), IAvH-Am-8850 (field number ARA 2348 ), collected by Andrés R. Acosta- Galvis and Carolina Huertas on February 15, 2004.
Type Locality. Tacarcuna farm, at the site La Gran Vía, kilometer 3 on the road to Cachipay, municipality of La Mesa, department of Cundinamarca, Colombia, 04°39’6.77’’ N, 74°25’1.0’’ W, 1300 m a.s.l.
Paratypes. ( Figures 5–6 View FIGURE 5 View FIGURE 6 ) Adult males IAvH-Am-8851, IAvH-Am-8853-4; adult female IAvH-Am-8855, MUJ-5002 ( ARA 2520 , GenBank accession numbers DQ502693 View Materials , DQ502261 View Materials , DQ502924 View Materials DQ503270 View Materials , DQ502394 View Materials , DQ503171 View Materials , DQ503396 View Materials , in Grant et al. 2006) same data as holotype and collected on October 10, 2004. Adult males IAvH-Am-8858-9, IAvH-Am-8862, adult females IAvH-Am-8863-5 from El Danubio farm, Viotá municipality, Cundinamarca Department, Colombia, 04°26’N, 74°28’ W, 1100 m a.s.l.; collected by Oscar Ibarra, Sandra González and Andrés Acosta on September 29, 2007. Adult females ICN-MHN-39106-07, adult male ICN-MHN 39105 from Torova creek , km 19 Cimitarra to school La Esperanza , Cimitarra municipality, vereda San Isidro and El Azufrado , Santander Department, Colombia, 06°22’60’’ N, 74°3’0’’ W, 565 m a.s.l., collected by Claudia Veléz on October 7, 1995. Adult male IAvH-Am-6554, from Chataná Creek , km 16 road to San Pablo of Borbur, Pauna municipality, Boyacá Department, Colombia, 05°40’35.6’’ N, 74°2’50.7’’ W, 1180 m a.s.l., collected by Mariela Osorno in 1999 GoogleMaps .
Referred specimens. (Juveniles and poorly preserved specimens)
Adult females, IAvH-Am-8852, IAvH-Am-8856; adult male IAvH-Am-8857, same data as holotype and col- lected on October10, 2004. Adult males IAvH-Am-8860-1, from El Danubio farm, Viotá municipality, Cundinamarca Departament, Colombia, 426’ N, 74 28 W, 1100 m a.s.l., MUJ-4451-2, MUJ-4454, MUJ-4456-57, MUJ-4459-64 from type locality. MUJ-3545-63, MUJ-3564 (GenBank accession numbers: DQ502610 View Materials , DQ502174 View Materials , DQ502877 View Materials , DQ503255 View Materials , DQ503370 View Materials , DQ503012 View Materials ), MUJ-3565-72 from La Magnolia farm, El Cural creek, El Totumo, municipality of Ibagué, Tolima Departament, 4°22’59.69’’ N, 75°11’55.96’’ W, 1047 m a.s.l., collected by Marco Rada and Santiago Sánchez on February 15, 2004 and November10, 2004 GoogleMaps .
Definition. A small species of Hyloxalus characterized by: (1) SVL of adult males 14.6–16.7 mm. (15.4 0.7, n =12), adult females 16.0–17.0 mm. (16.5 0.4, n =5); (2) disc on Finger III not expanded, 1.25–1.4 times the width of the adjacent phalanx; (3) when adpressed, Finger I equal or slightly longer than Finger II; (4) fringes absent on the Fingers; (5) Finger III not swollen in males; (6) adult males with pale arm gland ( Figure 2 View FIGURE 2 G–H); (7) disc on Toe IV slightly expanded, 1.2–1.5 times the width of the adjacent phalanx; (8) fringe absent on toes; (9) outer tarsal fold absent; inner tarsal fold present, proximally as oval or elongated tubercle-like; (10) toes unwebbed; (11) penultimate phalanx of finger and toes with a small white spot dorsally in life; (12) dorsolateral stripe absent; (13) oblique lateral stripe present, extending from groin to eye ( Figure 4 View FIGURE 4 ); (14) ventrolateral stripe absent (in preservative), but as a series of white spots on ventrolateral flanks in life ( Figure 4 View FIGURE 4 ); (15) throat, chest, and anterior belly cream, sexually dimorphic; dark brown in adult males by diffuse dark stippling and entirely cream in adult females ( Figure 4 View FIGURE 4 ); (16) testes white; (17) cloacal tubercles absent; (18) median lingual process absent; (19) advertisement call consisting of a series of pulsed notes with a dominant frequency between 4808-6387 Hz.
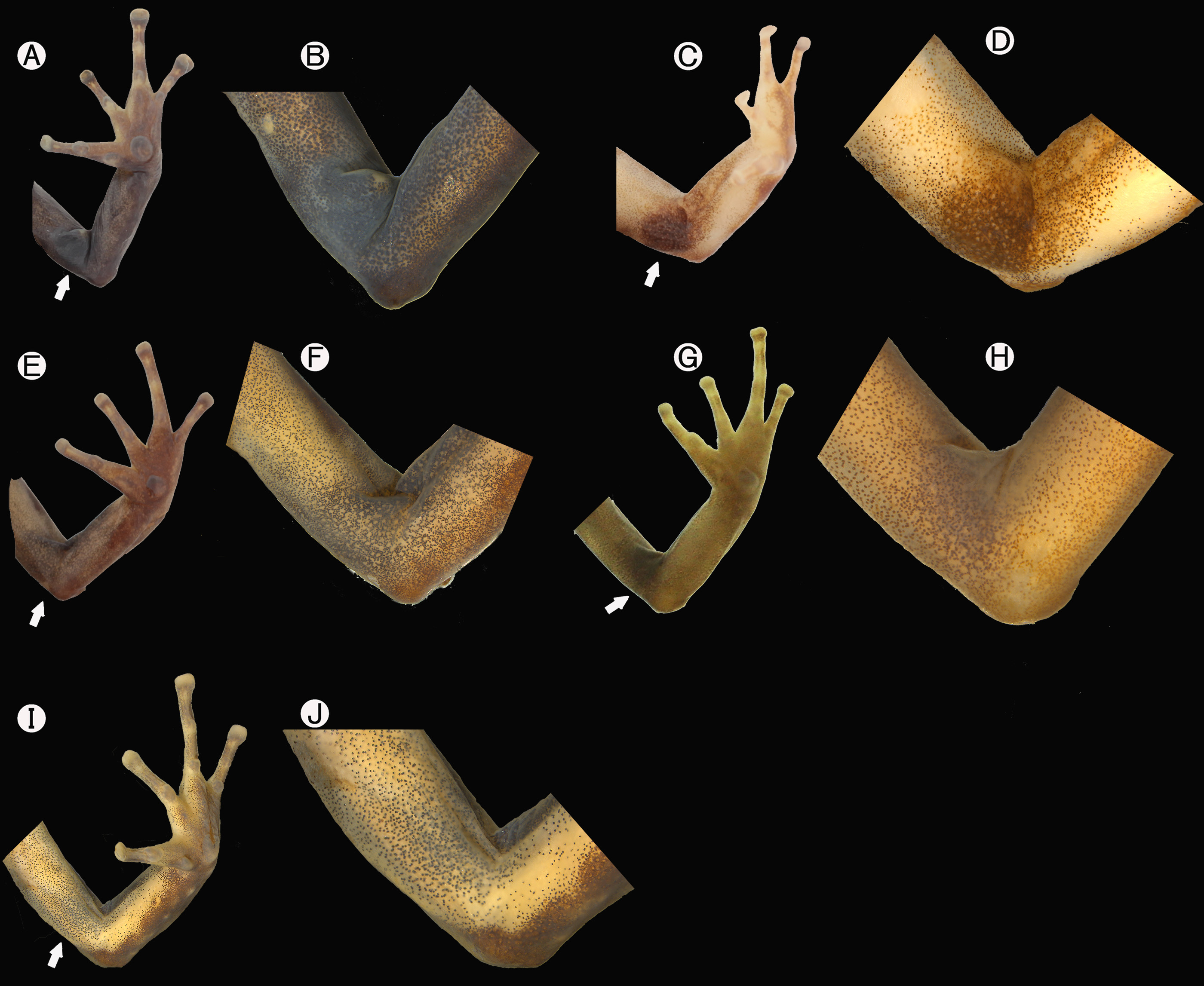
Diagnosis. Hyloxalus arliensis sp. nov. is a small frog ( Figures 5–6 View FIGURE 5 View FIGURE 6 ) with an arm gland in the adult males ( Figure 2 View FIGURE 2 ), a character which differentiates this species from other species of Hyloxalus , except for H. anthracinus , H. cevallosi , H. exasperatus , H. fascianigrus , H. lehmanni , H. ramosi , H. saltuarius , and H. nexipus (Grant & Castro- Herrera 1998; Grant & Ardila-Robayo 2002; Grant et al. 2006; 2017) ( Figure 7 View FIGURE 7 ).
Although the color of the arm gland in H. arliensis distinguishes the species from all Hyloxalus except for H. saltuarius , given the somewhat variable nature of gland color we provide comparisons against all Hyloxalus with arm gland, regardless of its color. Hyloxalus arliensis sp. nov. differs from H. fascianigrus by ventral coloration of the belly, cream with sparse stippling in adult males ( vs. large white spots on dark ground belly in H. fascianigrus ; Figure 7 View FIGURE 7 ), pale arm gland ( vs. black arm gland in H. fascianigrus ; Figure 2 View FIGURE 2 C-D), and 2–3 notes/ms (vs. 1 notes/ms in H. fascianigrus ). The new species differs from H. saltuarius by having the throat, chest, and anterior belly cream with diffuse dark stippling that darkens on males and is immaculate in females ( vs. completely black in both sexes in H. saltuarius ), and the complete lateral and solid oblique lateral stripe (OLS), (usually broken into a series of elongated spots in H. saltuarius ). The new species differs from Hyloxalus ramosi by having a dark longitudinal line on anterior surface of thigh (absent in H. ramosi ); the throat, chest, and anterior belly cream with diffuse dark stippling that darkening these regions in males vs. completely black in males of H. ramosi ; Figure 7 View FIGURE 7 ), pale arm gland ( vs. black arm gland in H. ramosi ; Figure 2 View FIGURE 2 A-B), and by the smaller size SVL in adult females (16.0–17.0 mm) and adult males (14.6–16.7) mm ( vs. SVL in adult females 20–20.2 mm and adult males 18.5–19.1 mm in H. ramosi ). Differs from H. exasperatus by having a complete oblique lateral stripe (incomplete, short, extending to 1/3 of the body in H. exasperatus sensu Coloma 1995 ), the absence of lateral keeled in toe IV (present in H. exasperatus sensu Coloma 1995 ), and lacking a dorsolateral stripe (present in H. exasperatus ). Differs from H. lehmanni by having smaller body size, SVL 14.6–16.7 mm in adult males and 16.0–17.0 mm in adult females ( vs. SVL 18.4–19.8 mm in adult males and 19.7–22.1 mm in adult females in H. lehmanni ), and the ventrolateral region of body without or with scarce white dots (notable white spots or wavy white ventrolateral stripe in H. lehmanni ). Differs from H. cevallosi by the absence of the ventrolateral stripe (present in H. cevallosi ), and having a smaller snout-vent length (SVL 14.6–16.7 mm) in adult males and (16.0–17.0 mm) in adult females ( vs. SVL 18.2 mm in adult male and 20.6–21.0 mm in adult females in H. cevallosi ). Hyloxalus arliensis sp. nov. differs from H. anthracinus by having a smaller size (SVL 14.6–16.7 mm) in adult males and (16.0–17.0 mm) in adult females ( vs. SVL 16.2–19.0 mm in adult males and 17.1–23.6 mm in adult females in H. anthracinus sensu Coloma 1995 ), and ventral coloration of the belly, cream with sparse stippling in adult males (solid black darkening on abdomen in males, venter white in females sensu Coloma 1995). Finally, H. arliensis sp. nov. differs from H. nexipus by lacking fringes on hands (a well-developed fringes in H. nexipus sensu Coloma 1995 ), dorsolateral stripe absent (present and incomplete in H. nexipus sensu Coloma 1995 ) and the call repertoire of Hyloxalus arliensis sp. nov. is composed of peep calls with a dominant frequency 4808–6387 Hz ( vs. trills calls with 2500–4700 Hz in H. nexipus sensu Coloma 1995 ).
Measurements of Holotype (in mm). SVL 16.5; forearm length 3.9; hand length 3.7; tibia length 7.4; foot length 7.3; head width 5.3; head length 7.0; eye diameter 2.0; eye to naris distance 1.6; internarial distance 2.2; snout length 2.3; interorbital distance 1.8; tympanum diameter 0.9.
Description and variation of the type series. Variation in meristic measurements are provided in Table 3. Adult males 14.6–16.7 mm ( n =12) SVL and adult females 16.0–17.0 mm ( n =5). Skin of dorsum, lateral and ventral surfaces smooth, posterior dorsum slightly granular, more notable in adult males (IAvH-Am-8851, IAvH-Am-8859- 60, IAvH-Am-8862); eyelids, posterior dorsum, thighs, and preaxillary area without tubercles, but the preaxillary surface with bulged aspect in some adult males (ICN-MHN-39105, IAvH-Am-8858) and tubercle-like ( Figure 4 View FIGURE 4 ). Cloacal tubercles absent. A small cloacal sheath covering cloaca (not as H. edwardsi species group, see discussion in Grant et al. 2006: 52). Males with unpigmented (white) testes.
Head width (between angles of jaws) 27.8–36.4 % of SVL, 0.9–1.1 times head length. Interorbital distance 30.7–46.9 % of head width. Snout sloped, bluntly rounded in dorsal aspect, sharply rounded in lateral view. Canthus rostralis well-defined and rounded. Loreal region flat or weakly concave, vertical, not sloping inward to lip ( Figure 4 View FIGURE 4 ). Eye diameter 33.8–44.5 % of head length. Eye-naris distance 40–79.3 % of snout length, 51–95 % of eye diameter. Nares slightly protuberant, directed posterodorsally. Anterior three-quarters of tympanum and annulus timpanicum partially visible, rest of tympanum concealed posterodorsally by relatively large; its diameter is 44.2–76.8 % of eye diameter. Low supratympanic bulge. Teeth present on maxillary arch.
Forelimbs moderate in length, forearm 19.6–25.4 % SVL. Hand length 23.3–28 % of SVL and 74.6–96.3 % of forearm length. Discs moderately expanded width of the adjacent phalanx 45.6–75.5 % of disc of Finger III. Finger III not swollen in adult males nor in adult females. Fringes absent on fingers. Finger I slightly longer than (usually) or subequal to Finger II when adpressed. Finger II reaches (ICN-MHN specimens) or extends beyond (usually) distal subarticular tubercle of Finger III; Finger IV beyond or reaches the distal (in ICN-MHN specimens) subarticular tubercle of Finger III. Relative lengths of adpressed fingers III>IV≥II≥I. Subarticular tubercles 1-1-2-2. Supernumerary tubercles absent; subarticular tubercles elliptical; thenar tubercle elliptical; palmar tubercle weakly subelliptical or subcircular; thenar tubercle from one-half to one third that of palmar tubercle; metacarpal fold or ridge absent ( Figure 8 View FIGURE 8 ).
Hindlimbs moderate in length, femur length 42.3–52.4 % of SVL and foot length 37–48.8% of SVL. Toe webbing and toe fringes completely absent. Relative lengths of adpressed toes IV>III>V>II>I. Disc of Toe III barely beyond the penultimate subarticular tubercle of the Toe IV; Toe V extended beyond the level of penultimate subarticular tubercle of Toe IV ( Figure 5 View FIGURE 5 ). Discs of the toes moderately expanded, width of the adjacent phalanx 50.0–75.6% of disc of toe IV. Tibia length 42–52% of SVL. Number of subarticular tubercles: I = 1, II = 1, III = 2, IV = 3, V = 2. Supernumerary tubercles absent; inner metatarsal tubercle weakly elliptical; outer metatarsal tubercle subcircular, smaller than inner metatarsal tubercle. Diameter of outer metatarsal tubercle 56.1–90.9% of inner metatarsal tubercle, outer tarsal fold absent; tarsal keel subcircular and elongate in shape of tubercle-like, diagonal to the tarsus, but without ridge nor fold, and not extended to pre-axial side of Toe I; without median metatarsal tubercle ( Figure 8 View FIGURE 8 ).
Color in preservative (ethanol 70 %, Figures 4 View FIGURE 4 and 6 View FIGURE 6 ). Dorsal surface brown to blackish-brown, sometimes with a black dorsal X-mark on dorsum ( Figure 6 View FIGURE 6 ). Pale dorsolateral stripe absent; upper eyelid region blackish brown in dorsal view; dorsal surfaces of thigh and of the hind limb with diffuse transversal dark brown stripes. A conspicuous brown facial stripe that extends from under eye to the groin; tympanum dark brown and cream; oblique lateral stripe cream and complete. Ventrolateral region of the body and lip cream. Anterior surface of thigh cream with a longitudinal stripe diffuse brown and posterior surface cream with diffuse dark brown motted or reticulated. Ventral surface of hindlimbs cream; plantar surfaces brown with diffuse cream mottling. Ventral region cream with diffuse brown melanophores; the throat, chest and part of anterior belly sexually dimorphic, darker with concentrated melanophores in adult males and immaculate white in adult females ( Figure 6 View FIGURE 6 ).
Color in life ( Figure 5 View FIGURE 5 ). Based on field notes deposited at MUJ and IAvH-Am. Dorsal view, light brown with copper iridescence and some black spots or with a design in X-shape on dorsum; forelimbs light brown; dorsal surfaces of thighs light brown, with black or brown transverse bands; anterior thigh cream or yellow with a brown longitudinal band; axillary and groin region yellow cream. Flanks of the head with a black band that extends from the anterior tip of the snout to the posterior region of forearm. Ventrolateral flanks of the body with white spots from the groin to axilla ( Figure 5 View FIGURE 5 ); upper and lower lip cream; an iridescent cream longitudinal stripe on infraorbital region that extends from anterior eye to under tympanum; iris copper with black reticulations. Ventral surfaces of the throat cream with scattered black melanophores reaching chest height in adult males; in adult females, the throat is cream to yellow; belly cream with some widely scattered black melanophores. Oblique lateral stripe cream with gold spots. Ventral surface of the thigh at the cloacal region with scattered and small white and black melanophores; palmar and plantar surfaces brown.
Etymology. The specific epithet refers to the ancient name of the Magdalena River; used by the indigenous Carares who lived in the middle basin of the Magdalena River, until the first half of the 20th century. The word arli means river of fish.
Distribution. Hyloxalus arliensis sp. nov. occurs at elevations between 565 and 1200 m a.s.l. on Andean slopes from the Magdalena valley in Antioquia, Boyacá, Cundinamarca, Santander and Tolima departments. Currently known from six localities ( Figure 9 View FIGURE 9 ), five of these localities lie on the western slope of the Cordillera Oriental of Colombia ( Figure 9 View FIGURE 9 A–E). The other sixth locality lies on the eastern slope of the Cordillera Central ( Figure 9F View FIGURE 9 ).
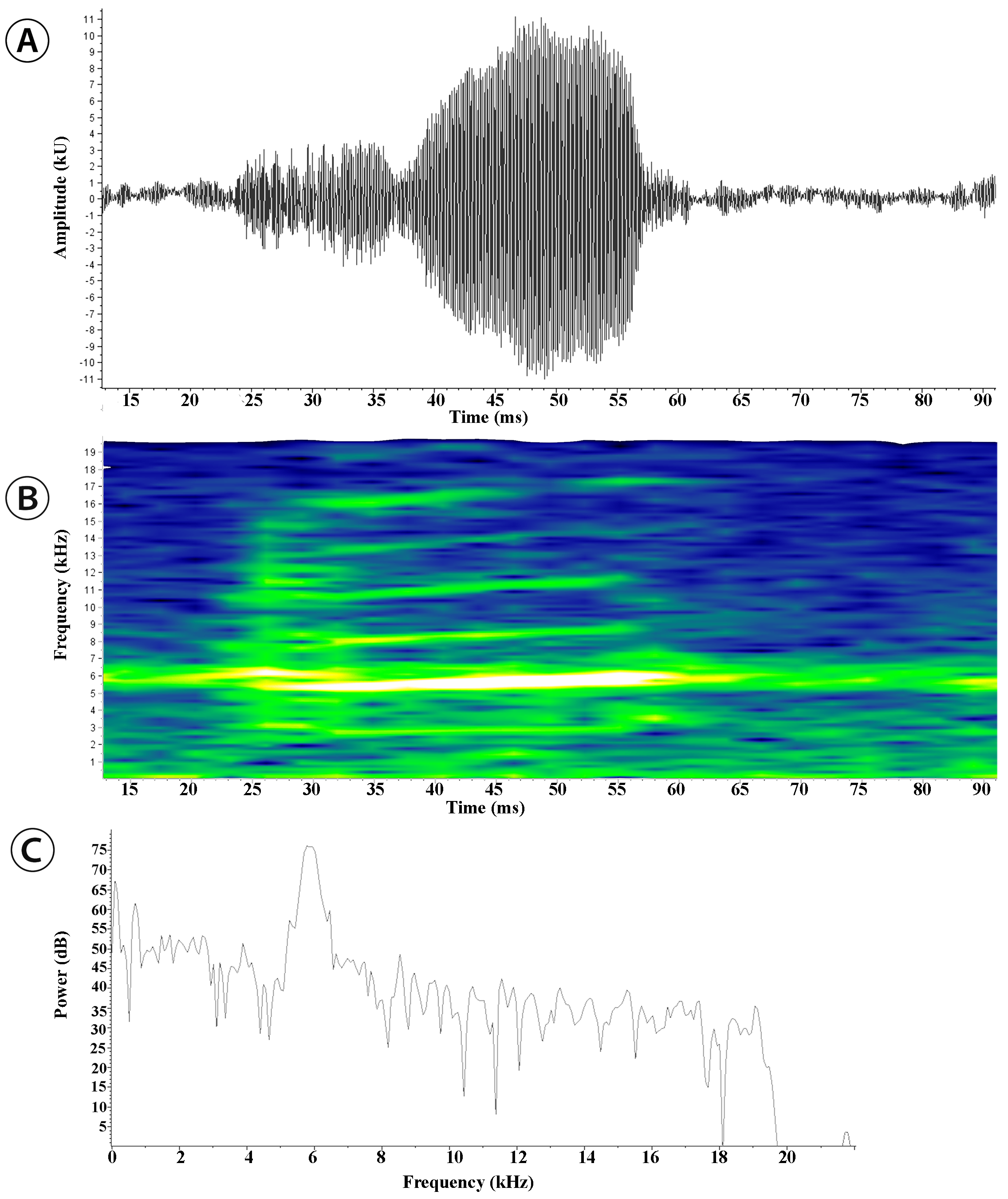
Advertisement call. Hyloxalus arliensis sp. nov. was recorded vocalizing hidden under leaf litter or ferns at the base of bushes near slow-flowing streams ( Figures 10–11 View FIGURE 10 View FIGURE 11 ), up to 1 m distant from the riverside, active during the rainy season. We recorded two minutes of advertisement calls on November 15, 2007 in the municipality of Viotá, Cundinamarca Department, Colombia ( 4°26’ N, 74°25’ W, datum WGS 84, 1100 m a.s.l.). Two specimens (IAvH- Am-8861-2) were recorded (BSA-15993) between 16:25–16:40 hours at a temperature ranging from 28–28.9 oC and at an environmental relative humidity between 64–65%. Temporal structure: The advertisement call of Hyloxalus arliensis sp. nov. consists of long trains of single notes (“peep”) with rapid and successive repetitions (two to three notes per milisecond) ( Figure 10 View FIGURE 10 ). The note rate varied between 89–132 notes/min. The note duration ( Figure 11 View FIGURE 11 ) was between 0.022 – 0.041 s (mean 0.035 + 0.005, n = 23), with 198–223 (mean 209 + 7.31, n = 22) oscillations contained in each note. The internote interval varied between 0.322 – 0.638 s (mean 0.42 + 0.006, n = 22). Spectral structure: Five to six harmonics were developed, with the dominant frequency 4808–6387 Hz (mean 5635 + 717, n = 25) and fundamental frequency in 2266–4086 Hz (mean 3037 + 787, n = 21).
Natural History. Hyloxalus arliensis sp. nov. is a diurnal species inhabiting heterogeneous agricultural areas ( Figure 12 View FIGURE 12 ) in the lowland Andes of the middle Magdalena River valley, associated with vegetation that borders slow flowing streams. Some specimens were found up to four meters away from the creek, in deforested areas dominated by grasslands or shrubby vegetation. This species was active during the rainy season between February to May and September to November. Generally, adult males vocalize after rains, calling from the ground hidden at the base of plants, usually Araceas or ferns; juvenile and post-metamorphic individuals were found hidden in the leaf litter. The clutch size consists of eight pigmented eggs ( Figure 12 E View FIGURE 12 ). Tadpoles are located in pools separate from the streamflow. Hyloxalus arliensis sp. nov. is syntopic with other species of the superfamily Dendrobatoidea ( sensu Grant et al. 2006) such as Rheobates palmatus and Dendrobates truncatus .
Remarks. Based on the observations and records obtained directly from fieldwork on two localities (Viotá and La Mesa Municipalities from the years 2004 to 2007) Hyloxalus arliensis sp. nov. was an abundant species with explosive breeding, highly seasonal (very active during rainy season) and occupying areas seriously affected by anthropogenic disturbance. Their relative abundances in this locality, indicate that they have adapted well to this disturbance. Their original habitats in the tropical Andean forest are subject to extensive conversion by agricultural use and the expanding human settlements producing habitat degradation and loss. With regard to the specific microhabitats of Hyloxalus arliensis sp. nov. (small streams), it is remarkable that they show such an association given the interruption of natural flows due to water management infrastructure, the presence of solid waste, and chemical pollution by agrochemicals. The natural condition and population dynamics in other localities are unknown and currently, there are not known records in protected areas including its original natural environment. Additionally, the available records of H. arliensis sp. nov. reveal an extension of 223 km in a straight line from its extreme localities (Ibaguée-Tolima to Cimitarra-Santander), which allows categorization as a Least Concern (LC) species (IUCN Red List, 2014).








| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.