
Imogine qeshmensis, Maghsoudlou, Abdolvahab & Momtazi, Farzaneh, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3753.4.5 |
|
publication LSID |
lsid:zoobank.org:pub:4556045E-C319-4549-995A-814011D9B684 |
|
DOI |
https://doi.org/10.5281/zenodo.6129855 |
|
persistent identifier |
https://treatment.plazi.org/id/8E8539C5-CC88-4929-89BA-9B746A49B3F1 |
|
taxon LSID |
lsid:zoobank.org:act:8E8539C5-CC88-4929-89BA-9B746A49B3F1 |
|
treatment provided by |
Plazi |
|
scientific name |
Imogine qeshmensis |
| status |
sp. nov. |
Imogine qeshmensis View in CoL sp. nov.
( Figures 3–4 View FIGURE 3 View FIGURE 4 )
Material and examined. Six specimens were collected in February 2011: two mature specimens provided serial sagittal sections, to elucidate the histological structure of the reproductive systems; one specimen was used for a whole mount as an entire animal stained with Carmine Alum. The remaining three specimens were preserved in ethanol 70%.
Holotype: one mature specimen, ZUTC Platy 1265 HS, with the anterior half preserved in ETOH 70%, and part of the posterior half containing the copulatory apparatus as series of sagittal sections (eight slides).
Paratypes: Five specimens: one sectioned specimen, ZUTC Platy 1266 HS, with the anterior half preserved in ETOH 70 %, and part of the posterior half containing the copulatory apparatus as series of sagittal sections (seven slides); one entire specimen, ZUTC Platy 1267 WM, cleared and stained with Carmine Alum; three entire worms, ZUTC Platy 1268 S, preserved in ETOH 70%.
Etymology. The specific epithet refers to Qeshm Island, where the specimens were collected.
Type locality. All specimens were collected from the upper intertidal rocky shore of Hamun jetty beach, Qeshm Island, Persian Gulf, Iran, 26° 58´35.3"N, 56° 14´53" E ( Fig. 1 View FIGURE 1 ). Habitat included rocky-muddy substrate ( Fig. 2 View FIGURE 2 A) covered with rock oysters ( Saccostrea cucullata ), other associated animals with the specimens included sponges and red tunicates.
Diagnosis. Body thick, fleshy and broadly oval; rosy brown with darker brown spots, become smaller toward margin, body margin free of spots with white opaque band. With small white knobbed tentacles. Tentacular, cerebral and frontal eyes present. Marginal eyes encircle the whole body margin. Without spermiducal bulb, ejaculatory duct joins short prostatic duct in middle of penis. Penis papilla small; male and female antra ciliated. Gonopores well separated.
Type of the genus: Imogine oculifera Girard, 1853 .
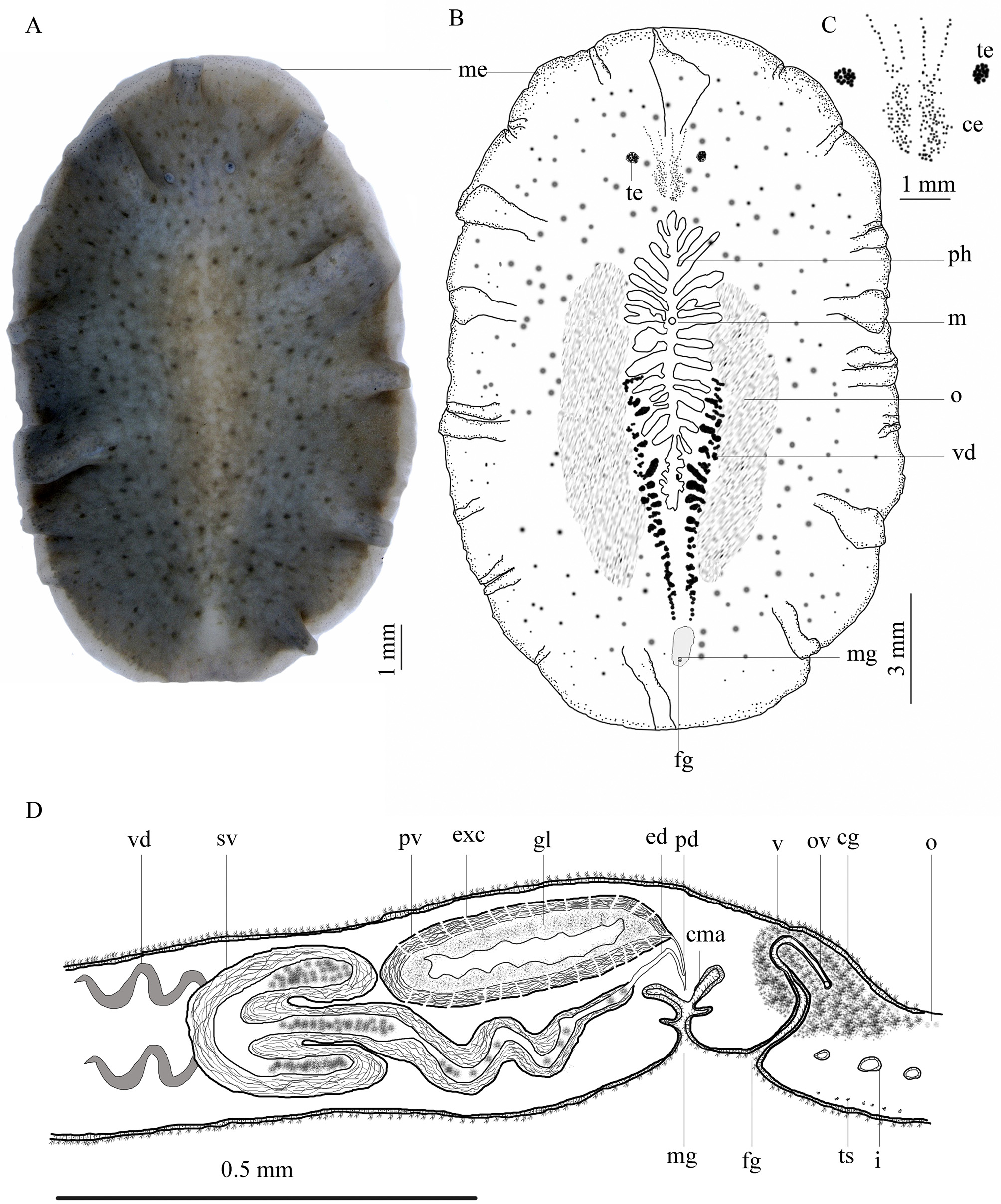
Description. External morphology. Body broadly oval, thick and fleshy with deep marginal ruffles ( Fig. 3 View FIGURE 3 A). Holotype 25 × 18 in size, the average size of paratypes 17 × 12. Dorsal body color of living animals rosy brown, lighter in the middle of the body around the pharynx ( Fig. 2 View FIGURE 2 B), with regular dark brown spots, become smaller toward margin; body margin with wide white opaque band, 0.5 mm in thickness, free of spots ( Fig. 3 View FIGURE 3 A), covered with eye spots ( Fig. 3 View FIGURE 3 A, B). More or less translucent cream, in ventral view, vas deferens visible on eider sides of pharynx, as two dark, cream, coiled strips, extends anteriorly from the male gonopore to middle half of body ( Fig. 3 View FIGURE 3 B). Nuchal tentacles small, rounded, retractile and white, 2.2 apart; tentacular eyes with 18–21 large eye spots at tip of each tentacle ( Fig. 3 View FIGURE 3 C); small cerebral eyes embedded in epidermis between nuchal tentacles, difficult to observe, but readily observable on cleared anterior part of holotype and stained whole paratype (ZUTC Platy.1267 WM), cerebral eye clusters more or less paired, more crowded posterior to nuchal tentacles with 55 eye spots in each posterior cluster, cerebral eyes extend anterior to nuchal tentacles; frontal eyes, with about 21 eyes in each cluster, scattered and subdivided ( Fig. 3 View FIGURE 3 C), frontal eyes not reaching anterior margin. Marginal eyes extend along the entire margin, denser anteriorly, in four or five rows, reducing posteriorly into two or three rows ( Fig. 3 View FIGURE 3 B).
Ruffled pharynx located centrally, 4.7 from the anterior end, 7.3 from posterior end of the body, 12.5 mm in length, with about 12 pairs of lateral folds, mouth located in anterior part of middle third of pharynx ( Fig. 3 View FIGURE 3 B). Male pore located posterior to pharynx, 3.8 apart ( Fig. 3 View FIGURE 3 D).
Epidermis and body wall. Ventral epidermis three times thicker than dorsal epidermis, 95–100 µm thick ( Figs. 4 View FIGURE 4 B, D). Cellular epidermis consists of thick-walled (18.5 µm) columnar cells and rhabdites, basal membrane 6.7 µm in thickness; arrangement of muscle fibers beneath basement membrane: outer, very thin circular muscle layer (3.6 µm), followed internally by thick longitudinal fibers (11.5 µm), then well-developed diagonal muscles (31 µm) and innermost longitudinal muscles merging with diagonal muscle fibers (26 µm). Transverse muscle fibers well developed ( Fig. 4 View FIGURE 4 B).
Dorsal epidermis about 36 µm thick ( Fig. 4 View FIGURE 4 A); cellular epidermis consists of tall, thin-walled columnar cells, ciliated, with intra-epithelial nuclei and rhabdites; basal membrane about 7 µm in thickness; body wall musculature consists of outer, very thin circular layer, followed by inner longitudinal layer (12 µm). Clusters of granular pigment scattered beneath dorsal and ventral wall throughout the parenchyma; few sparse rhabdites observed below dorsal epidermis ( Fig. 4 View FIGURE 4 A).
Reproductive structures. Male reproductive system with tripartite seminal vesicle, free, elongated oval prostatic vesicle, without spermiducal bulb, and penis papilla housed in a ciliated antrum ( Figs. 3 View FIGURE 3 D & 4C). Testes scattered ventrally throughout body length. In ventral view, vasa deferentia arising in mid-body, converging posteriorly on either side of pharynx ( Fig. 3 View FIGURE 3 B). In sagittal view, vas deferens recurrent below prostatic vesicle, opens into lateral lobes of seminal vesicle ( Fig. 3 View FIGURE 3 D). Seminal vesicle 620 µm wide, enclosed in strong muscular layer (100 µm in thickness), located anterior to prostatic vesicle, oriented horizontally in holotype ( Fig. 4 View FIGURE 4 C) but, oriented slightly dorsally in sectioned paratype. Lateral lobes of seminal vesicle equal in size, 860 µm long × 200 µm wide, central lobe larger than lateral lobes, 1.2 mm long × 200 µm wide ( Figs. 3 View FIGURE 3 D & 4C). Central lobe tapers posteriorly below the prostatic vesicle to form straight ejaculatory duct, near distal end of prostatic vesicle; ejaculatory duct joins short prostatic duct in middle of penis. Prostatic vesicle surrounded by muscular layer, 80 µm in thickness, 1 mm long × 400 µm width ( Figs. 3 View FIGURE 3 D & 4C). Wall of prostatic vesicle with numerous narrow regular transverse ducts (extra-vesicular gland canals) leading into lumen, with polyglandular lining, glandular cell-bodies of extra-vesicular glands localized more or less distant in parenchyma surrounding male apparatus ( Fig. 4 View FIGURE 4 D). Penis papilla small, 170 × 250 µm, housed in ciliated, deep and narrow male antrum ( Fig. 3 View FIGURE 3 D).
Female gonopore 230 µm posterior to male gonopore; vagina narrow, somewhat muscular and curved, with cuboidal epithelium, vagina proceeds dorsally, and then curves backward to receive uteri; vagina surrounded by numerous cement glands, with ciliated female antrum ( Fig. 3 View FIGURE 3 D). Lang's vesicle absent. Male and female gonopores separated ( Fig. 3 View FIGURE 3 D).
Remarks. Faubel (1983), in his classification, emended the family Stylochidae and introduced the main diagnostic character of the family as: the ridged glandular lining of the prostatic vesicle with extra-vesicular glands. Recently, Bulnes et al. (2005) defined the family Stylochidae based on two new types of prostatic linings: the polyglandular and the mono-glandular types. In the case of mono-glandular type each extra-vesicular gland is connected via a glandular duct with a single inner tube, so the number of extra-vesicular glands and inner tubes is equal. In poly-glandular type, each long-fingered extension is connected via several glandular ducts to extravesicular glands ( Bulnes et al. 2005). However, extra vesicular glands were not observed by Newman and Cannon (1996a, b) in some members of the family including Imogine and Stylochus .
Based on the presence / absence of a tripartite seminal vesicle, Marcus and Marcus (1968) divided the genus Stylochus within the Stylochidae , into the two subgenera: Imogine Girard, 1853 and Stylochus Ehrenberg, 1831 . Prudhoe (1989) and Newman et al. (1993) recognized 28 species of Stylochus within the subgenus Imogine . Later, Jennings and Newman (1996b) elevated the Imogine to generic rank. Then, several new species were assigned to this genus around the world ( Bulnes 2010; Jennings & Newman 1996a; Lee 2006; Sluys et al. 2005).
The genus Imogine is defined by a tripartite and highly muscular seminal vesicle and a prostatic vesicle of the polyglandular type ( Bulnes et al. 2005). Within the genus, the species are discriminated by their color pattern, body shape and consistency, presence or absence of tentacles, the number and pattern of tentacular and cerebral eyes, and the presence of frontal and marginal eyes. Due to the phenotypic variability of these characters, additional features such as the position of pharynx and mouth, the presence of spermiducal bulbs, the position of the junction between the ejaculatory duct and the prostatic duct, the form and size of the penis papilla and the histology of the male atrium have been considered ( Bulnes 2010).
The presence of a tripartite seminal vesicle clearly places the specimen described here as a member of the genus Imogine . Currently, this genus contains 36 species ( Bolaños 2010) with two taxonomic groups within the species: those with eyes around the entire body margin and those with only anterior eyes ( Newman et al. 1993). Sixteen of them, like the species describe here, have eyes around the entire body margin (Table 1). Jennings and Newman (1996a) have summarized the main diagnostic characters of Imogine species with eyes around the body margin. These characters with some modifications are listed in Table 1.
In terms of geographical distribution (Table 1), the aforementioned sixteen species occur in the following regions: Atlantic Ocean (eight species), Mediterranean Sea (one species), north Pacific (one species) and Indo West Pacific (six species). Jennings and Newman (1996 a, b) considered the geographical distribution as a character to discriminate their five new species of Imogine inhabiting the Indo-West Pacific from all other congeners with eyes around the entire body margin. We believe that history and intensity of the maritime activities in the Persian Gulf provides the possibility (through discharge of ballast water) to introduce species from other parts of world’s ocean into the region. Therefore, we compared our species from the Persian Gulf with all of the above mentioned sixteen species.
Based on the information given in Table 1, and compared with the Imogine species inhabiting the Atlantic Ocean, the species described here is distinguished from: (1) Imogine arenosus by its color pattern, tentacle color and number of tentacular eyes. I. arenosus is dorsally reddish brown with white and black dots, has numerous tentacular eyes at the base of tentacles, while the species described here is dorsally rosy brown with regular dark brown spots, these spots become smaller towards the margin; the nuchal tentacles are small, retractile and white in the fixed specimens; with 18–21 tentacular eyes at the tip of each tentacle (2) I. catus in terms of color pattern, shape of tentacles, shape of the prostatic vesicle and body size. The body in I. catus dorsally has brown spots in two lateral bands, unlike the species described here. The species presented here is 25 × 18 in size, with small nuchal tentacles, and elongated oval prostatic, while I. catus is 65 × 32 mm in size with elongated tentacles and a spherical prostatic vesicle (3) The presence of cilia in the male and female antra of the species described here distinguishes this species form I. meridianus that lacks ciliated male and female antra ( Bulnes 2010); in the present species the ejaculatory duct joins the prostatic duct in the middle of the penis, while in I. meridianus the ejaculatory duct joins the prostatic duct at the proximal end of the penis, (4) I. minimus differs from the species described here in terms of body size, color pattern, and size of the prostatic vesicle with respect to the seminal vesicle. The body of I. minimus is relatively small, 3 × 3 mm, the dorsal surface is light brown with dark grey flecks and brown spots, and the prostatic vesicle is twice the size of the seminal vesicle, while in the species described here seminal vesicle is equal in size with the prostatic vesicle; (5) I. pulcher differs from the species presented here in terms of color pattern, numerous tentacular and frontal eyes, and marginal eyes few and scattered in the posterior half of the body, while in the present species tentacular eyes numbered 18–21 per tentacle, with about 40 frontal eye spots, and marginal eyes extending well posteriorly as two visible rows. I. pulcher has a pyriform male antrum and an extremely small penis papilla, while in our species the male antrum is deep and narrow, and the penis papilla is not extremely small (170 µm long × 250 µm wide); (6) I. refertus has a dorsal surface which is brown with a bright orange mesh, few cerebral eyes that differ in number and distribution from our species; (7) I. zebra is clearly different from the species presented here in terms of color pattern, number of tentacular eyes, frontal eyes extending to the anterior margin, having a spherical and erect prostatic vesicle, while the prostatic vesicle in the species described here is elongated, oval and horizontal; (8) I. mediterraneus is elongated and oval unlike the species presented here that is broadly oval in form, the cerebral eyes number 50 eyes in each cluster, the prostatic vesicle is nearly twice the size of the seminal vesicle, the ejaculatory duct joins distally to the prostatic duct, while in our species the cerebral eyes numbered 91 eyes; the prostatic vesicle is equal in size to the seminal vesicle and the ejaculatory duct joins the prostatic duct in the middle of the penis papilla; (8) I. oculiferus , as type of the genus, is dorsally cream to brown with red or pink spots, with a moderately thick muscular coat and a pyriform prostatic vesicle, an ejaculatory duct which joins the prostatic duct at the base of penis, and with a spermiducal bulb ( Hyman 1940), while the species described here has an elongated oval and highly musculature prostatic vesicle,, and lacks a spermiducal bulb.
Compared with the Indo-West Pacific species of Imogine , the species described here is distinguished from all other congeners except I. pardalotus by: having cerebral eyes as two parallel clusters, while the Indo-West Pacific species described by Jennings and Newman (1996 a, b) have a cerebral eyes as single cluster. The number of tentacular eyes, 18–21 per tentacle, of the species described here is considerably lower than all of the species of Imogine from the Indo-West Pacific. As indicated in Table 1, the number of tentacular eyes per tentacle is: 100 eyes for I. kimae , 50 eyes for I. lesteri , 30 eyes for I. mcgrathi , 50 eyes for I. meganae and 100 eyes for I. pardalotus . I . mcgrathi also differs from the species described here by having numerous frontal eyes.
I. pardalotus , unlike the species described here, has an elongated oval body, 15 × 6 mm, the dorsal color is beige with a leopard spotted pattern of greenish brown spots, irregular in shape and color, with elongate tentacles and the ejaculatory duct joins the prostatic duct at the proximal end of penis papilla (Table 1). I. meganae has yellow margins, with elongate yellow tentacles, and is clearly distinguished from the species described here that has no marginal band and small, retractile, white tentacles. I. kimae has a prostatic vesicle twice the size of the seminal vesicle and the ejaculatory duct joins the prostatic duct at the proximal end of the penis papilla, unlike our new species.
Based on the Figures 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 of Jennings and Newman (1996a) for I. kimae , I. mcgrathi , I. meganae and I. pardalotus , the seminal vesicle is located ventrally to the prostatic vesicle, while in the species described here the seminal vesicle is entirely anterior to the prostatic vesicle ( Figs. 3 View FIGURE 3 D, 4C).
The ejaculatory duct joins the prostatic duct at the proximal end of the penis papilla in I. lesteri , the situation which is not similar to our species. I. exigus is relatively smaller (7 × 4 mm) than present species (25 × 18 mm). I. exigus also has fewer cerebral eyes (7 eyes) compared with the species described here (about 91 eyes). Based on the above diagnostic features, Imogine qeshmensis is a new species.
In terms of habitat, members of the genus Imogine are known to be associated with or feed on giant clams and rock oysters ( Jennings & Newman 1996a; Newman et al. 1993; Sluys et al. 2005), or other invertebrates such as barnacles (Lee et al. 2006). All specimens of the species described here associated with rock oysters. Our sampling area located at Qeshm Island is a free economic zone. The island is used for mariculture such as cultivation of shellfish and pearl oysters. The presence of the new species described here should be considered by managers of oyster farms because these flatworms kill and consume cultured bivalves, significantly contributing to mortalities ( Galleni et al. 1980; Littlewood & Marsbe 1990; Newman et al. 1993).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.