
Troglotayosicus meijdeni, Botero-Trujillo, Ricardo, González-Gómez, Julio C., Valenzuela-Rojas, Juan C. & García, Luis F., 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4244.4.7 |
|
publication LSID |
lsid:zoobank.org:pub:3A7B3322-6F2D-417B-9774-C5BA8FF67325 |
|
DOI |
https://doi.org/10.5281/zenodo.5683196 |
|
persistent identifier |
https://treatment.plazi.org/id/03A587D4-2B25-FFB6-A9AB-697FFBD3FA86 |
|
treatment provided by |
Plazi |
|
scientific name |
Troglotayosicus meijdeni |
| status |
sp. nov. |
Troglotayosicus meijdeni sp. nov.
Figures 1–9 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 ; Table 1 View TABLE 1
Type material. Holotype. Female from COLOMBIA: Huila : Rivera, Vereda Agua Caliente, area adjacent to Termales Los Ángeles , 880 m, 02°45′06.6"N 75°14′17.0"W, i.2014, J.C. González Gómez, J.C. Valenzuela Rojas (MPUJ-ENT 0046833). Paratypes GoogleMaps . Same data of holotype: 1 female (AMNH), 1 female (ICN-AS-1001), 1 female (MACN-Ar), 1 female (MACN-Ar), 1 juvenile male (MPUJ-ENT 0046834), 1 juvenile female (ICN-AS-1002).
Etymology. The species is named for Dr. Arie van der Meijden, who participated in the expedition that discovered this Troglotayosicus population. JCGG, JCVR and LFG would like to honor Van der Meijden for his collaboration with the BEA Group, to which they belong, in the study of Colombian biodiversity. Van der Meijden conducts research on scorpion biomechanics at CIBIO Research Centre in Biodiversity and Genetic Resources, InBIO, Universidade do Porto, Portugal.
Diagnosis (female-based only). The new species differs from others in the genus by: (i) lateral eyespot large, being the same size of the PDMi or larger ( Fig. 3 View FIGURE 3 B, Table 1 View TABLE 1 ); (ii) pedipalp chela DMA and DI carinae coarsely granular anteriorly ( Figs. 6 View FIGURE 6 A, D); (iii) metasomal segments III, IV ventral surface smooth, without any trace of VL or VSM carinae ( Figs. 8 View FIGURE 8 B–C); (iv) telson vesicle markedly inflated (telson L:vesicle W ratio 2.13:1 to 2.18:1; telson L:vesicle H ratio 3.20:1 to 3.26:1), as compared to that of T. hirsutus and T. humiculum , such that the apex of the aculeus does not reach the level of the vesicle ventral margin in lateral aspect ( Fig. 3 View FIGURE 3 G); and (v) pedipalp chela noticeably incrassate ( Figs. 2 View FIGURE 2 E, 6), e.g., chela L:H ratio equal to 2.40:1.
Comparisons. Females of the other known Troglotayosicus species differ from the above description by: (i) lateral eyespot small, its diameter less than half that of the PDMi, in T. hirsutus and T. humiculum , whereas it is not clear if the eyespot is present or absent in T. vachoni ; (ii) pedipalp chela DMA and DI carinae identified by subtle differences in angles between adjacent surfaces (as other carinae), without granulations, or DMA and DI barely granular, in T. hirsutus , whereas the chela of T. vachoni was described as smooth, without carinae or granulations; (iii) metasomal segments III, IV ventral surface granulose, with VL and VSM carinae, in T. humiculum , whereas some granules are reportedly present on ventral posterior half of segment IV, in T. vachoni ; (iv) telson vesicle moderately inflated or elongated in shape, at least in T. hirsutus (telson L:vesicle W ratio 2.18:1 to 2.21:1; telson L:vesicle H ratio 3.16:1 to 3.24:1) and T. humiculum (telson L:vesicle W ratio 2.45:1 to 2.64:1; telson L:vesicle H ratio 3.26:1 to 3.60:1), while the apex of the aculeus reaches (or slightly surpasses) the level of the vesicle ventral margin in lateral aspect; and (v) the pedipalp chela of females of the three species is narrower than that of the female of the new species; e.g., chela L:H ratio 2.89:1 to 3.13: 1 in T. humiculum (according to Ochoa et al. 2010), 3.09:1 to 3.31: 1 in T. hirsutus (according to Botero-Trujillo et al. 2012), and 3.47: 1 in T. vachoni (according to Lourenço 1981).
Female holotype Female paratype (MPUJ-ENT 0046833) (ICN-AS-1001)
Total body L:1 27.25 29.60
Carapace: L: 3.59 3.99 Lateral ocelli:7 PLMa / PDMi diameter: 0.15 / 0.08 0.17 / 0.10
Eyespot diameter: 0.10 0.10 Description. Based on five females and two juveniles. Measurements in Table 1 View TABLE 1 .
Color. Figs. 2 View FIGURE 2 C–E, 3, 9. Description from preserved specimens. Base color, chelicerae and legs yellowish to chestnut; carapace, tergites, metasoma, and pedipalps yellowish (juveniles) or carapace, tergites and pedipalps chestnut, darker on pedipalps and metasomal segments (adults); sternites, coxosternal region and legs yellowish, darker in adults; pectines whitish. Carapace and coxosternal region with array of yellowish marks, lighter than base color; pedipalp manus (more in adults) with longitudinal darker areas in the position of the carinae. PLMa and PDMi ocelli surrounded by black pigment; eyespot unpigmented.
Chelicerae. Fig. 4 View FIGURE 4 . Manus, dorsal and external surfaces predominantly smooth, sparsely setose; ventral surface densely setose. Fixed finger, dorsal margin with four teeth (distal, subdistal, median, and basal); median and basal teeth separate, not fused into bicusp. Movable finger, internal (dorsal) margin with five teeth [distal, two small subdistal, median, and basal]; internal distal and external (ventral) distal teeth not opposable, internal distal tooth at most partially overlapping external distal tooth in dorsal view; ventral surface with long, well-developed serrula, starting at level of internal basal tooth and reaching slightly beyond internal distal tooth.
Carapace. Figs. 3 View FIGURE 3 A–B, 5A. Anterior margin sublinear, with weak median projection (epistome) and bilaterally-symmetrical array of macrosetae; posterior margin sublinear, with few macrosetae. Median eyes absent. Type 2B pattern of lateral ocelli: two ocelli (PLMa, PDMi), ALMa, MLMa, ADMi, PLMi absent ( Loria & Prendini 2014); eyespot present; PLMa largest, about one and a half to almost twice the diameter of the PDMi ocellus and of the eyespot, both of which are subequal in size. Median longitudinal sulcus evident but shallow, not reaching anterior margin; posterolateral and posterior transverse sulci obsolete. Surfaces acarinate and smooth, with some setae.
Pedipalps. Figs. 6 View FIGURE 6 , 7 View FIGURE 7 . Carinae well developed (adults and juveniles). Femur tricarinate; DI, DE and VI carinae complete, with coarse granules; VE carina absent; dorsal intercarinal surface with scattered granules; internal surface with abundant prominent granules; ventral and external surfaces smooth; all surfaces with macro- and microsetae. Patella bicarinate; DI and VI carinae complete and coarsely granular; all other carinae absent; DPP consisting of three prominent granules subproximally; VPP absent; intercarinal surfaces smooth, with macro- and microsetae. Chela manus markedly incrassate, such that chela height 1.04 to 1.07 times greater than fixed finger length, and chela L:H ratio equal to 2.40:1. Chela with DMA and DI carinae coarsely granular on distal third and distal half of manus, respectively; VE carina smooth but pronounced; DS, D, E and VI carinae identified by subtle differences in angles between adjacent surfaces and without granulations. Manus, intercarinal surfaces smooth, covered with macro- and microsetae, mostly along the carinae, and becoming more numerous on fingers. Fixed finger, median denticle row comprising six oblique, slightly imbricate primary subrows of denticles, flanked by five external and six internal denticles; terminal denticle considerably larger than preceding denticles, hook-like; fingertips interlocking unevenly when closed such that movable finger displaced to exterior. Movable finger, median denticle row comprising seven oblique, slightly imbricate primary subrows of denticles, flanked by six external and seven internal denticles; terminal denticle enlarged, hook-like, accommodated in subdistal diastema of fixed finger.
Trichobothria. Figs. 6 View FIGURE 6 , 7 View FIGURE 7 . Femur with three trichobothria: one external (e), one dorsal (d), one internal (i). Patella with 19 trichobothria, two petite (et2, esb2), one accessory (em3): two ventral (v1, v2); 14 external (et1–et3, est, em1–em3, esb1, esb2, eb1–eb5); two dorsal (d1, d2); one internal (i). Chela with 26 trichobothria, five petite (Et4, Et5, Esb, Db, dsb), one accessory (dm): 15 on manus, four ventral (V1–V4), ten external (Et1–Et5, Est, Esb, Eb1–Eb3), one dorsal (Db); 11 on fixed finger, four external (et, est, esb, eb), five dorsal (dt, dst, dm, dsb, db), two internal (it, ib).
Legs. Legs I–IV, all segments with setae variable in length. Femur with rows of granules along dorsal (absent in leg I) and ventral edges. Patella with VE carina made up of small granules on distal two-thirds of legs I, II, obsolete in III, IV; other carinae absent. Remaining segments of legs acarinate and smooth. Tibial spurs absent; prolateral pedal spurs present; retrolateral pedal spurs vestigial. Basitarsi I–IV setose, each (except IV) with series of brush-like spinules distally; dorsal retrolateral and median prolateral brushes well developed on legs I–III; ventral prolateral brush vestigial on legs I, II, apparently absent (probably obsolete) on III; ventral retrolateral brush obsolete to vestigial on legs I, II, apparently absent on III. Telotarsi I–IV, dorsomedian lobe with one large seta; ventral surface without spinules, subspiniform setae regularly arranged into pair of parallel ventrosubmedian rows; ungues well developed, curved, equal in length; dactyl shorter than ungues.
Tergites. Fig. 5 View FIGURE 5 C. Pretergites smooth. Posttergites I–VI, surfaces predominantly smooth (especially in juveniles), finely granular along posterior margin; I–VII with bilaterally-symmetrical array of macrosetae.
Posttergite VII, surface finely granular, posterior half with scattered granules in position of DSM carinae; DL carinae present in posterior half, each terminating in tubercle comprising two or three granules; posterior margin coarsely granular.
Sternum. Fig. 5 View FIGURE 5 B. Shape subpentagonal with rounded apex; posterior width greater than length; lateral margins sub-parallel, converging anteriorly; posterior depression shallow. Surface with 10 or 11 macrosetae and additional microsetae.
Genital operculum and pectines. Fig. 5 View FIGURE 5 B. Genital operculum divided longitudinally, formed by subtriangular plates; pectinal plate, wider than long; surface of both with macro- and microsetae. Pectines, marginal lamella with the standard three segments; middle lamella comprising one large basal segment and an additional rounded segment. Tooth count 7– 7 in all females (n = 10 pectines) and juveniles (n = 4 pectines); proximal and (especially) distal teeth slightly larger than others. Lamellae and teeth with abundant macrosetae.
Sternites. Fig. 5 View FIGURE 5 D. Sternites III–VII, surfaces acarinate and smooth, each with abundant macro- and microsetae throughout and on lateral and posterior margins; III–VI each with pair of small, rounded spiracles, situated mediolaterally.
Metasoma. Segments I–V progressively increasing in length, with fairly abundant macro- and microsetae. Segments I–IV, dorsal intercarinal surfaces smooth; lateral intercarinal surface with sparse conspicuous granulation; ventral surfaces of segments I–IV smooth. Segment V, dorsal intercarinal surface smooth; lateral and ventral surfaces densely and coarsely granular, except on basal third were predominantly smooth; granules increasing in size posteriorly. DL carinae complete, costate-granular, converging distally on segments I–IV, complete, granular on V; LSM carinae vestigial, comprising row (or patch) of granules between DL and ML carinae, in medial third of segments I–IV, and in posterior two-thirds of V; ML carinae complete, costate-granular on segments I–IV, restricted to posterior two-thirds of V where granular; LIM carinae evident, comprising incomplete row of granules (somewhat disaggregated on IV) in posterior half (or posterior two-thirds) of segments I–IV, undistinguishable on V; VL and VSM carinae completely absent on segments I–IV; VL, VSM and VM carinae present in distal two-thirds of segment V, consisting of coarse granules mixed with scattered granulation.
Telson. Fig. 3 View FIGURE 3 G. Vesicle inflated, not much elongated; anterodorsal lateral lobes present; dorsal and dorsolateral surfaces smooth, ventral surface with abundant shallow granules arranged in three broad longitudinal areas; lateral and ventral surfaces with several macro- and microsetae. Aculeus moderately long, gently curved; apex not reaching the level of the vesicle ventral margin in lateral aspect.
Male. Unknown.
Distribution and ecology. Troglotayosicus meijdeni sp. nov. is known only from the type locality at Rivera municipality, Huila department, Colombia ( Fig. 1 View FIGURE 1 ). Specimens were collected at the border of a rudimentary local road ( Figs. 2 View FIGURE 2 A–B). All were extracted, upon ultraviolet light detection ( Stahnke 1972), from the leaf litter which had accumulated at the base of trees and shrubs ( Figs. 2 View FIGURE 2 C–D). Careful, manual scrutiny of the litter layer was the only search method, which proved useful for the detection of this species, whereas no individuals were found wandering on the surface.
Behavioral observations. One event of cannibalism between conspecific specimens of T. meijdeni sp. nov. was observed in the field. This involved a larger scorpion preying on another of smaller size. As documented in Fig. 9 View FIGURE 9 , the predator had the prey’s metasoma immobilized while stinging through a soft membrane, and also during consumption when the latter was still alive.
Some specimens of T. meijdeni sp. nov. were found immobile inside of the leaf litter layer. These individuals had the metasoma turned over the body, and the appendages (legs and pedipalps) tucked-in against the body. After remaining still for a few seconds, the animals turned the body and rolled into the substrate (“tumbling-in”). This behavior was observed in juvenile specimens; in contrast, adult females were more prone to attempt escaping upon perturbation of the substrate, although they adopted the same position when captured.
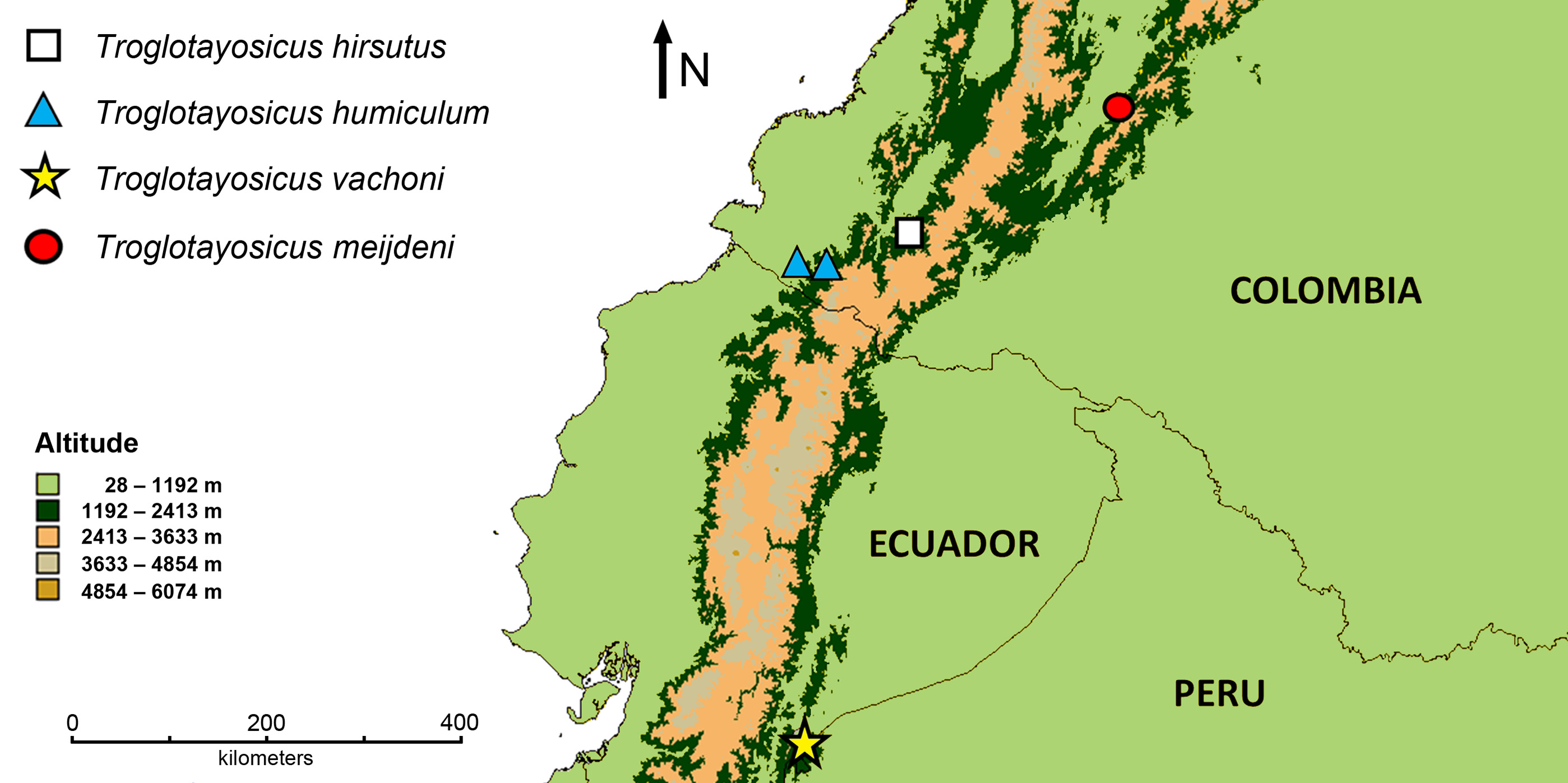


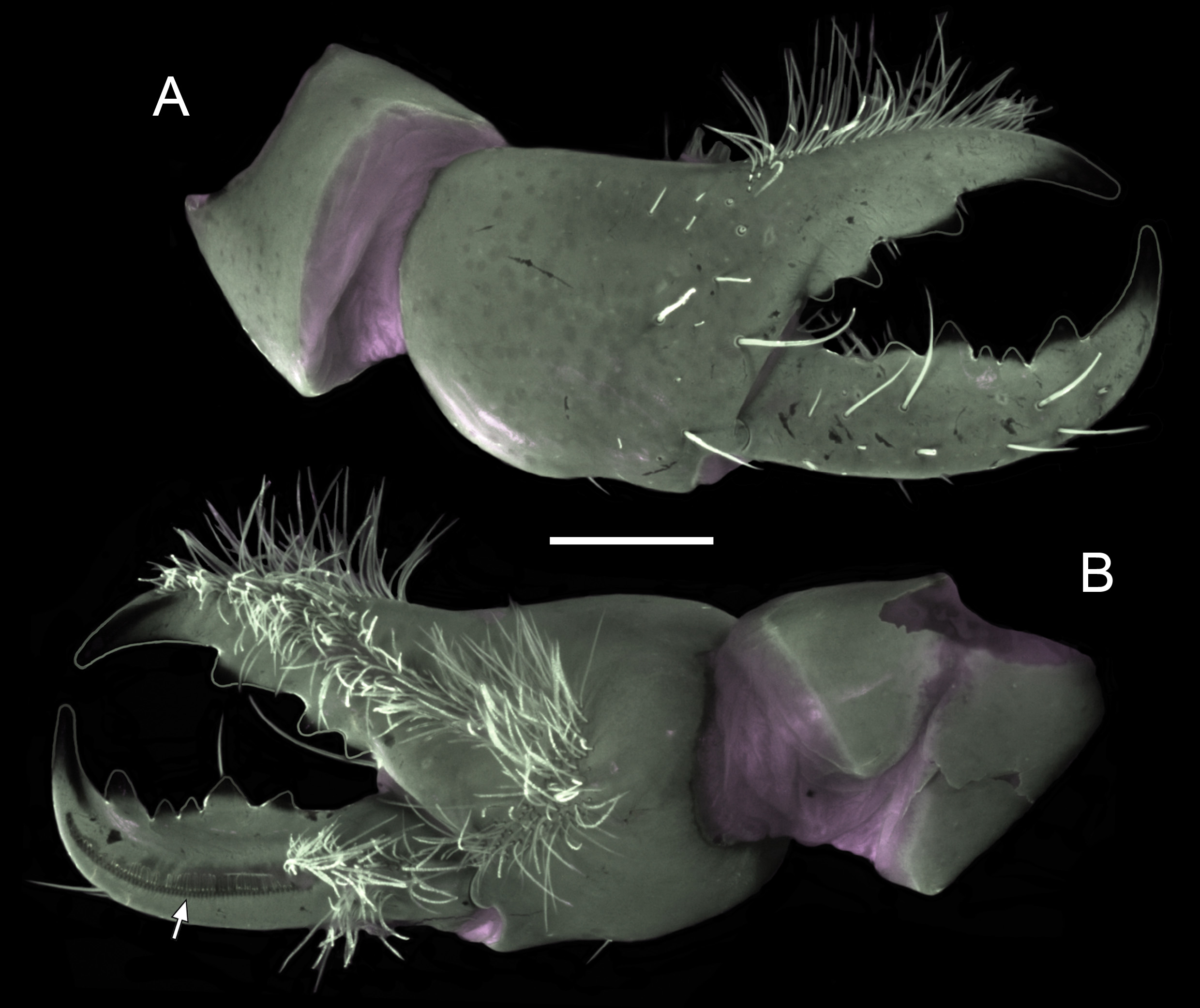

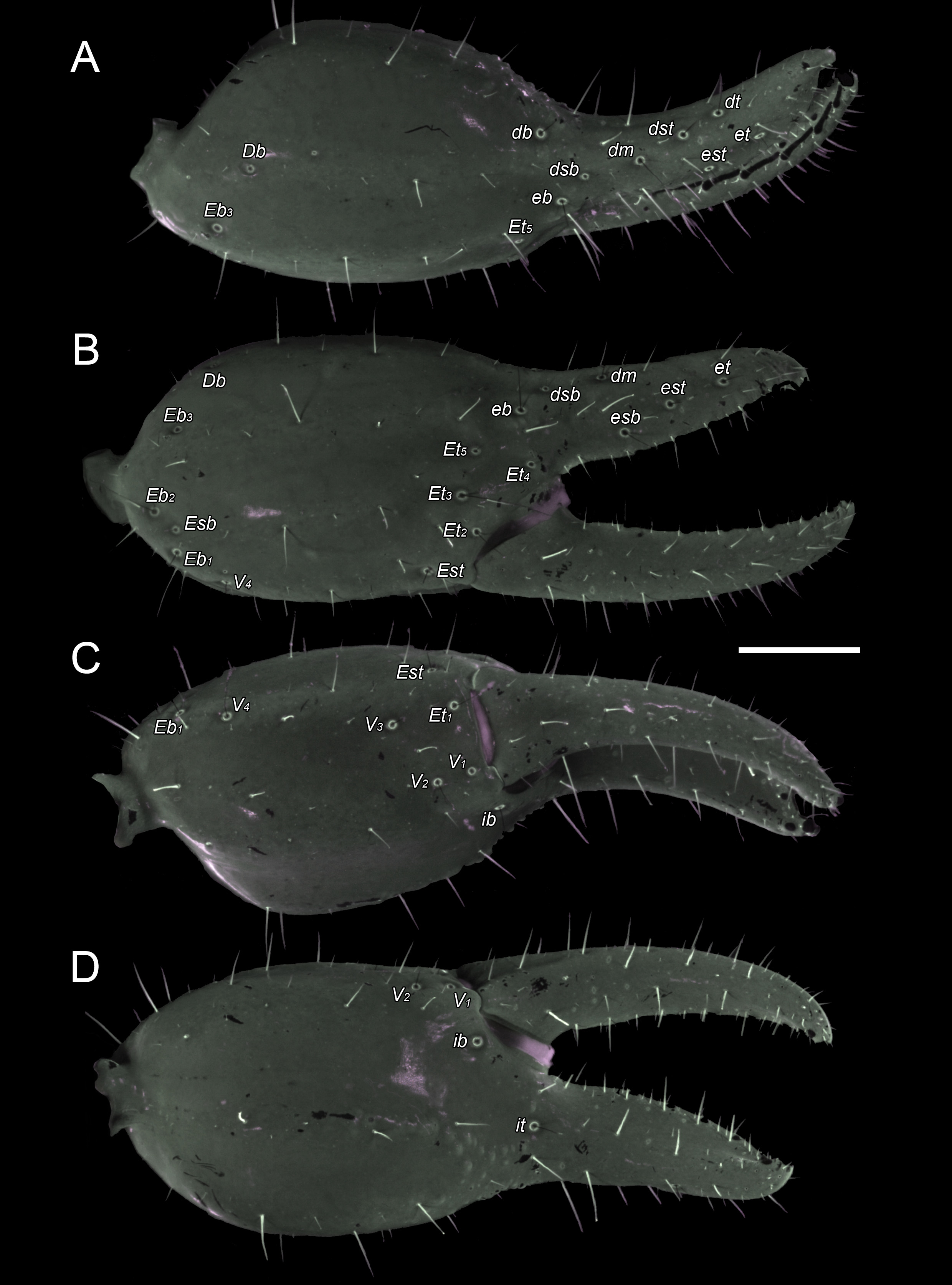

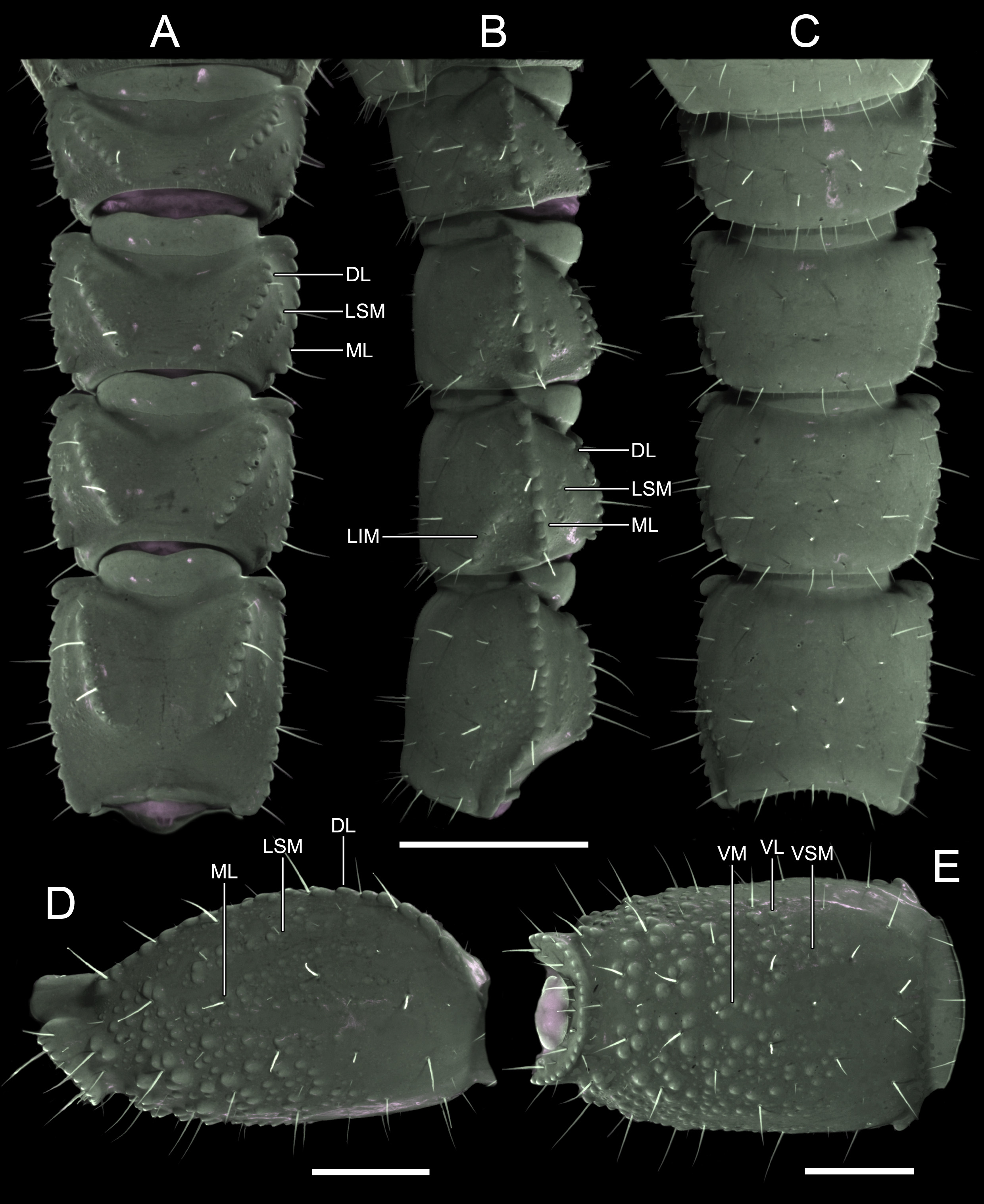

TABLE 1. Troglotayosicus meijdeni sp. nov. Metric data for two adult females. Measurements in millimeters. 1 Sum of carapace, mesosoma and metasoma. 2 Sum of pre- and postttergites I – VII, measured along the midline. 3 Sum of metasomal segments I – V and telson. 4 Sum of femur, patella and chela. 5 Measured from the commissure of the junction with the movable finger to the finger tip. 6 Measured along the marginal lamella. 7 Terminology for lateral ocelli follows Loria & Prendini (2014).
| Anterior W: Posterior W: | 2.39 3.59 | 2.53 3.86 | |
|---|---|---|---|
| Mesosoma: Metasoma: | Total L:2 Total L:3 | 7.93 15.73 | 8.07 17.54 |
| Metasomal segment I: | L: W: | 1.37 2.80 | 1.50 3.10 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |