
Teyuwasu barberenai Kischlat, 1999
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4629.1.12 |
|
persistent identifier |
https://treatment.plazi.org/id/03A6D706-FFCF-4E10-FF2D-FCA2FC20F8F5 |
|
treatment provided by |
Plazi |
|
scientific name |
Teyuwasu barberenai Kischlat, 1999 |
| status |
|
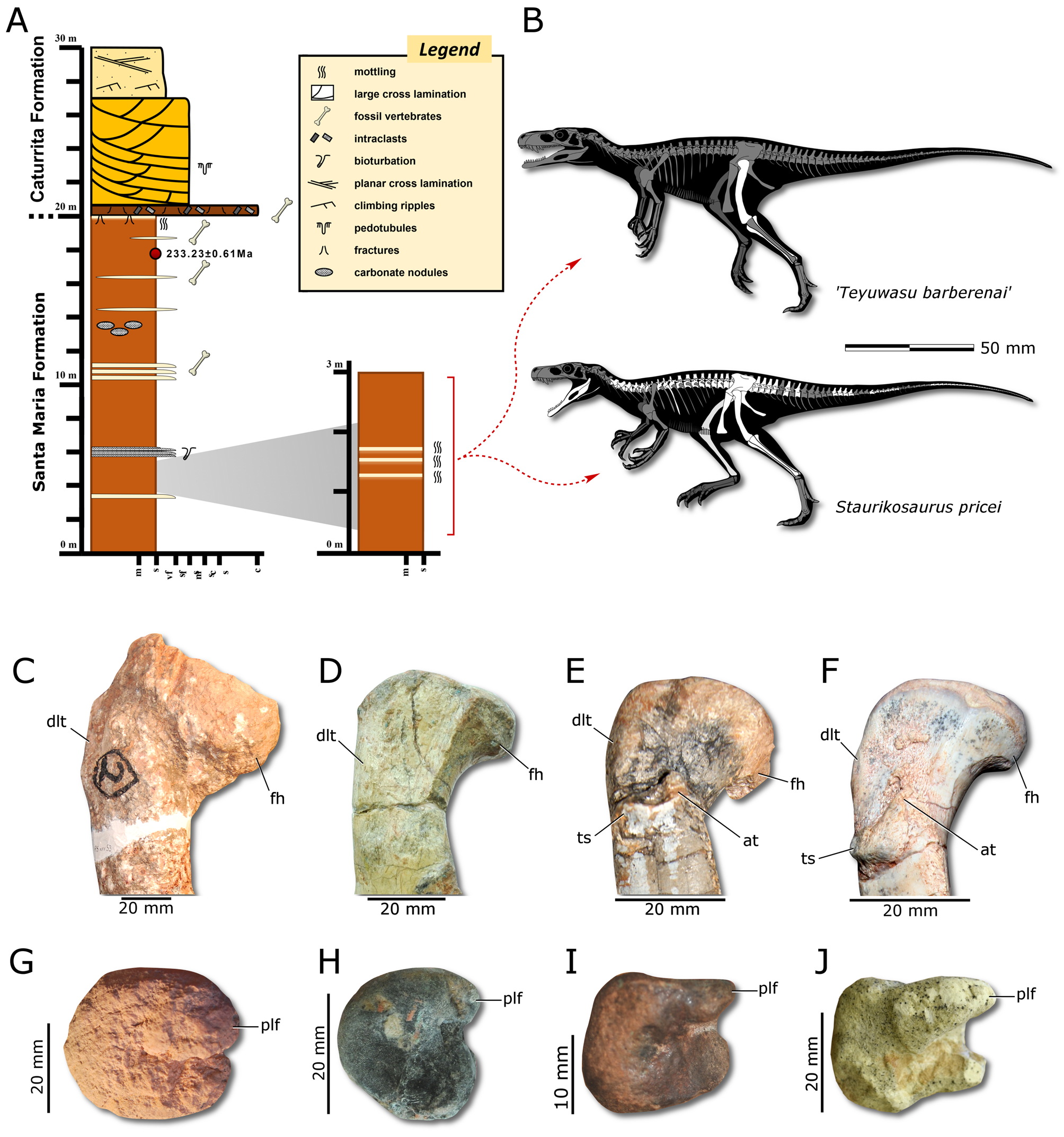
The controversial dinosauriform Teyuwasu barberenai Kischlat, 1999 ( Fig. 1B View FIGURE 1 ) is based on a right femur ( BSPG AS XXV 53 ) and tibia ( BSPG AS XXV 54 ), formerly referred to the pseudosuchian ‘ Hoplitosuchus raui ’ (= Hoplitosaurus raui ) Huene, 1938 (see also Huene, 1942). This material comes from a classic Late Triassic ( Carnian ) locality in southern Brazil ( Fig. 1A View FIGURE 1 ), the Cerro da Alemoa outcrop on the Alemoa complex, that has yielded several noteworthy tetrapod specimens (see Garcia et al., 2019, for a complete list of references) . When reviewing these abovementioned materials, Kischlat (1999) considered it to belong to a “robust saurischian dinosaur”, but later this taxon was considered to be a nomen dubium ( Langer et al., 2010; Ezcurra, 2012). Foremost, the initial description of this taxon is problematic, because Kischlat (1999) presented it in a symposium abstract which does not constitute a published work [which is not allowed under the Art. 9.10 of the International Commission on Zoological Nomenclature (ICZN)]. Moreover, the author did not list traits that clearly differentiated Teyuwasu from other coeval dinosauriforms (cf. Art. 13 of the ICZN, see further below). In particular, it was not adequately distinguished from Staurikosaurus pricei Colbert, 1970 ( Fig. 1B View FIGURE 1 ), which also comes from the same Alemoa complex, but from another nearby outcrop (Sanga Grande/Sanga de Baixo) considered equivalent in stratigraphic level and horizon with the lower levels of the Cerro da Alemoa site ( Huene, 1942; Colbert, 1970; see also Garcia et al., 2019) ( Fig. 1A View FIGURE 1 ). However, Huene (1942) did not clearly specify that the materials (femur and tibia) later assembled by Kischlat (1999) into the holotype of Teyuwasu were found in close association, although Huene (1942) cited that they come from the same stratigraphical level and horizon, and to our interpretation, their morphology and preservational features are compatible. Still, assigning these bones as part of a single individual is tentative.
In this contribution, we aimed to re-examine the holotype and only known specimen of Teyuwasu and reassess its validity and systematic position. Since Ezcurra (2012) recently described and compared the morphology of the materials attributed to this taxon, herein we focused on commenting only on the main features and characters pertinent to our systematic proposition. Likewise, our comparisons mostly encompassed dinosauriforms coeval to Teyuwasu (i.e., Carnian-earliest Norian). Firstly, as pointed by Ezcurra (2012), Teyuwasu can be recognized as a dinosauriform based on a combination of features: strongly inturned femoral head; longitudinal groove on the proximal surface of the femur; small posterior tuberosity of the femoral head; presence of an anterior trochanter on the femur; asymmetric posterior condyles of the proximal end of the tibia; tibia with a posterolateral process exceeding laterally the facet for the reception of the ascending process of the astragalus; tibial longitudinal lateral groove separating the posterolateral process and the facet for the reception of the ascending process of the astragalus.
Regarding morphological features, Kischlat (1999) stated that Teyuwasu lacked a femoral trochanteric shelf, and instead presented proximodistally directed parallel ridges of the homologous area. Whereas Teyuwasu indeed lacks a trochanteric shelf ( Fig. 1C View FIGURE 1 ), the condition of the aforementioned ridges is difficult to recognize because of the poor preservation of the specimen. Moreover, the trochanteric shelf is absent in Staurikosaurus as well, although Bittencourt & Kellner (2009) argued that a rugose surface is present on the homologous area of the left femur of the holotype ( Fig. 1D View FIGURE 1 ). Among mid to early Late Triassic dinosauriforms, a trochanteric shelf is absent in several groups, such as some ornithischians (e.g., Eocursor — Butler et al., 2007), theropods (e.g., Tawa— Nesbitt et al., 2009), and post-Carnian sauropodomorphs (e.g., Macrocollum— Müller et al., 2018). On the other hand, this feature is present in some non-dinosaurian dinosauriforms (e.g., Lewisuchus — Bittencourt et al., 2014 ), silesaurids (e.g., Silesaurus — Dzik, 2003 ), early sauropodomorphs (e.g., Bagualosaurus— Pretto et al., 2018; Buriolestes— Cabreira et al., 2016; Saturnalia— Langer, 2003) ( Figs. 1E, F View FIGURE 1 ), and herrerasaurids (Herrerasaurus— Novas, 1993; Sanjuansaurus— Martínez & Alcober, 2010). Therefore, the distribution of the trochanteric shelf regarding dinosauriforms of the initial Late Triassic is highly variable, though it is present in most taxa coeval to Teyuwasu . Following, the ventrally directed femoral head of Teyuwasu is not seen in Staurikosaurus and most basal dinosauriforms, though we interpreted this condition as result of taphonomic distortion, agreeing with Ezcurra (2012). Nevertheless, Teyuwasu exhibits a symmetric fourth trochanter, which is similar to the condition in nondinosaurian dinosauriforms (e.g., Lewisuchus , Silesaurus ), the herrerasaurid Staurikosaurus , some theropods (e.g., Coe-lophysis— Cope, 1889), but differs from most early sauropodomorphs (e.g., Buriolestes, Macrocollum , Saturnalia ), some herrerasaurids (e.g., Herrerasaurus , Sanjuansaurus ), some theropods (e.g., Tawa ), and ornithischians (e.g., Eocursor). Distally, the femur of Teyuwasu has a crista tibiofibularis that is not well separated from the lateral condyle by a groove, similar to the condition in herrerasaurids (e.g., Herrerasaurus and Staurikosaurus ), and on contrary of the condition in early sauropodomorphs (e.g., Buriolestes , Saturnalia ) and theropods (e.g., Tawa ). Moreover, the femoral size of Teyuwasu (c. 26 cm) is compatible with other herrerasaurids [ Staurikosaurus (c. 22 cm), Herrerasaurus (c. 36 cm on average) and Sanjuansaurus (c. 39 cm)], which are generally larger than other Carnian dinosauriforms [e.g., Saturnalia (c. 15 cm on average), Buriolestes (c. 14 cm), Bagualosaurus (c. 22 cm)].
In relation to the tibia, Teyuwasu has a distal end of the tibia with a rounded outline in distal view ( Fig. 1G View FIGURE 1 ), that contrasts with those of most coeval dinosauriforms, such as Bagualosaurus, Buriolestes , Herrerasaurus , and Saturnalia , which exhibit a subquadrangular or subretangular shape (Figs I, J). Indeed, a distal end of the tibia with a rounded outline in distal view was regarded as a possible autapomorphy of Staurikosaurus ( Bittencourt & Kellner, 2009) ( Fig. 1H View FIGURE 1 ), but was recognized in Tawa ( Nesbitt et al., 2009) and Sanjuansaurus ( Alcober & Martínez, 2010) . On the other hand, distinct from Tawa and other dinosauromorphs, the posterolateral flange of the distal end of the tibia of Teyuwasu does not exceed the lateral margin of the bone, same as in Herrerasaurus and Staurikosaurus ( Fig. 1G View FIGURE 1 ).
Additionally, using skeletal robustness of Teyuwasu to differentiate it from other coeval dinosauriforms is problematic because the holotypic bones are taphonomically expanded, as well as many of the tetrapod specimens that have been excavated from the Alemoa Member of the Santa Maria Formation (see Holz & Schultz, 1998 for more information). This is especially true for the specimens recovered from those distal floodplain horizons (see Garcia et al., 2019), so detection of this biased taphonomic trend depends on personal interpretation, which hinders additional morphological assignments.
As explained above, the anatomy of ‘ Teyuwasu’ differs from all other coeval dinosauriforms, except for Staurikosaurus . Indeed, the diagnosis proposed by Kischlat (1999) fails to distinguish ‘ Teyuwasu’ from the coeval Staurikosaurus , and accordingly, we did not recognize any unambiguous differences between both taxa, which share a unique combination of traits among coeval dinosauriforms: (i) femur without a trochanteric shelf; (ii) symmetric fourth trochanter of the femur; (iii) crista tibiofibularis poorly separated from the lateral condyle at the distal end of the femur; (iv) posterolateral flange of the distal end of the tibia of does not exceeds the lateral margin of the bone; (v) and rounded distal end of the tibia. Moreover, both taxa are similar in size and were exhumed from the same fossiliferous site complex. Except by Bagualosaurus, which differs anatomically from ‘ Teyuwasu’, no other described dinosauriform from the Santa Maria Formation reaches a similar size. Therefore, according to anatomical and geological data, we consider ‘ Teyuwasu barberenai’ a junior synonym of Staurikosaurus pricei , following the Principle of Priority of the ICZN (Art. 23). Since the description of Staurikosaurus by Colbert (1970), no other specimens had been referred to this taxon. Consequently, our proposal redefines BSPG AS XXV 53 and BSPG AS XXV 54 as the first specimens recognized as referable to Staurikosaurus for almost half a century.
Institutional abbreviations. BSPG, Bayerische Staatssammlung für Paläontologie und Geologie, Munich, Germany; CAPPA/UFSM, Centro de Apoio à Pesquisa Paleontológica da Quarta Colônia, Universidade Federal de Santa Maria, São João do Polêsine, Brazil; MCP, Museu de Ciências e Tecnologia, Porto Alegre, Brazil; MCZ, Museum of Comparative Zoology, Cambridge, USA; PVSJ, Instituto y Museo de Ciencias Naturales, San Juan, Argentina; ULBRA, Universidade Luterana do Brasil, Coleção de Paleovertebrados, Canoas, Brazil.
| BSPG |
BSPG |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.