
Chlorospingus flavovirens
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4136.2.7 |
|
publication LSID |
lsid:zoobank.org:pub:50C131BE-E262-4658-A53C-C11A1A709046 |
|
DOI |
https://doi.org/10.5281/zenodo.6056267 |
|
persistent identifier |
https://treatment.plazi.org/id/03A7326D-FFEF-8334-43F4-FC17FC52FC20 |
|
treatment provided by |
Plazi |
|
scientific name |
Chlorospingus flavovirens |
| status |
|
Results and discussion View in CoL
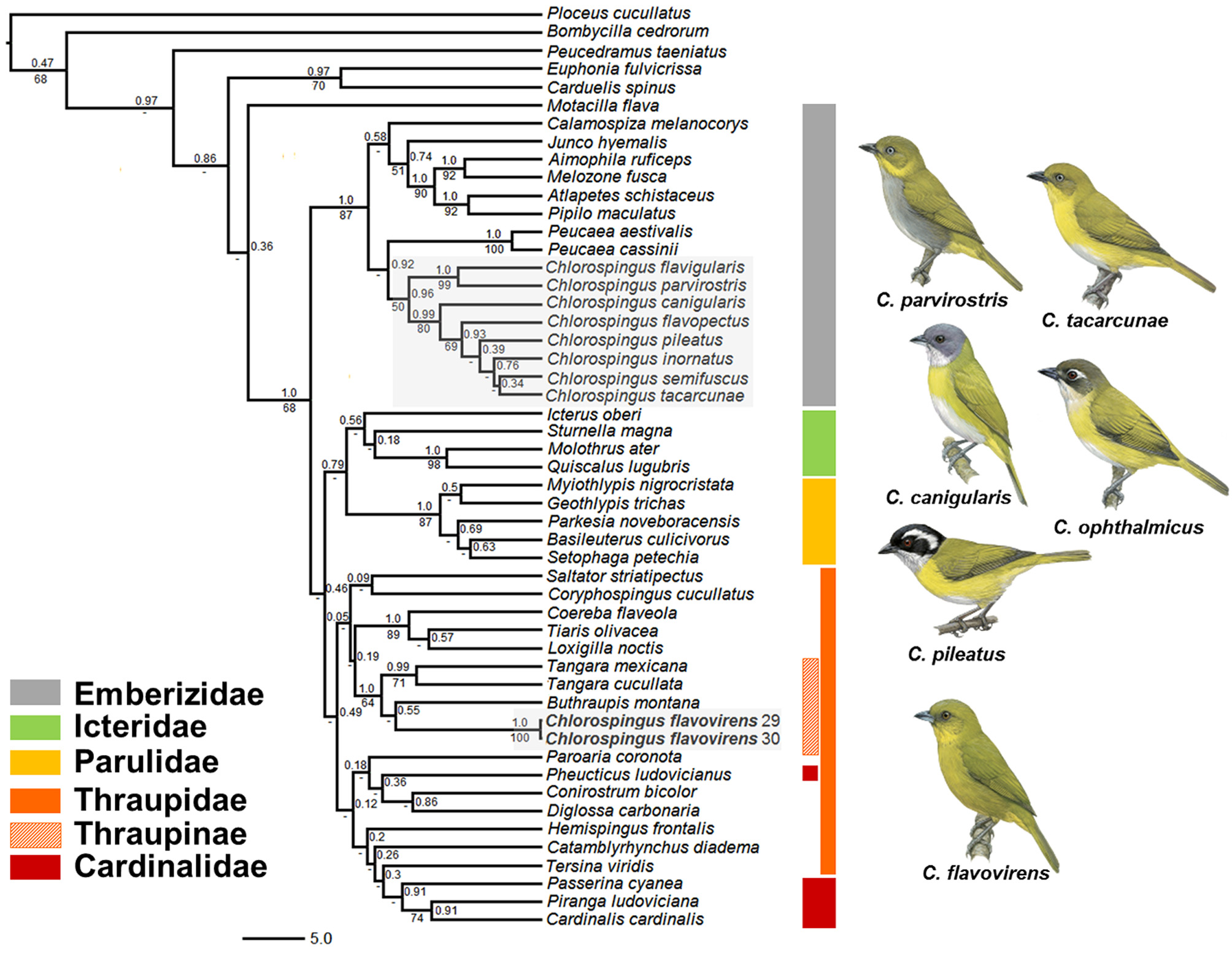
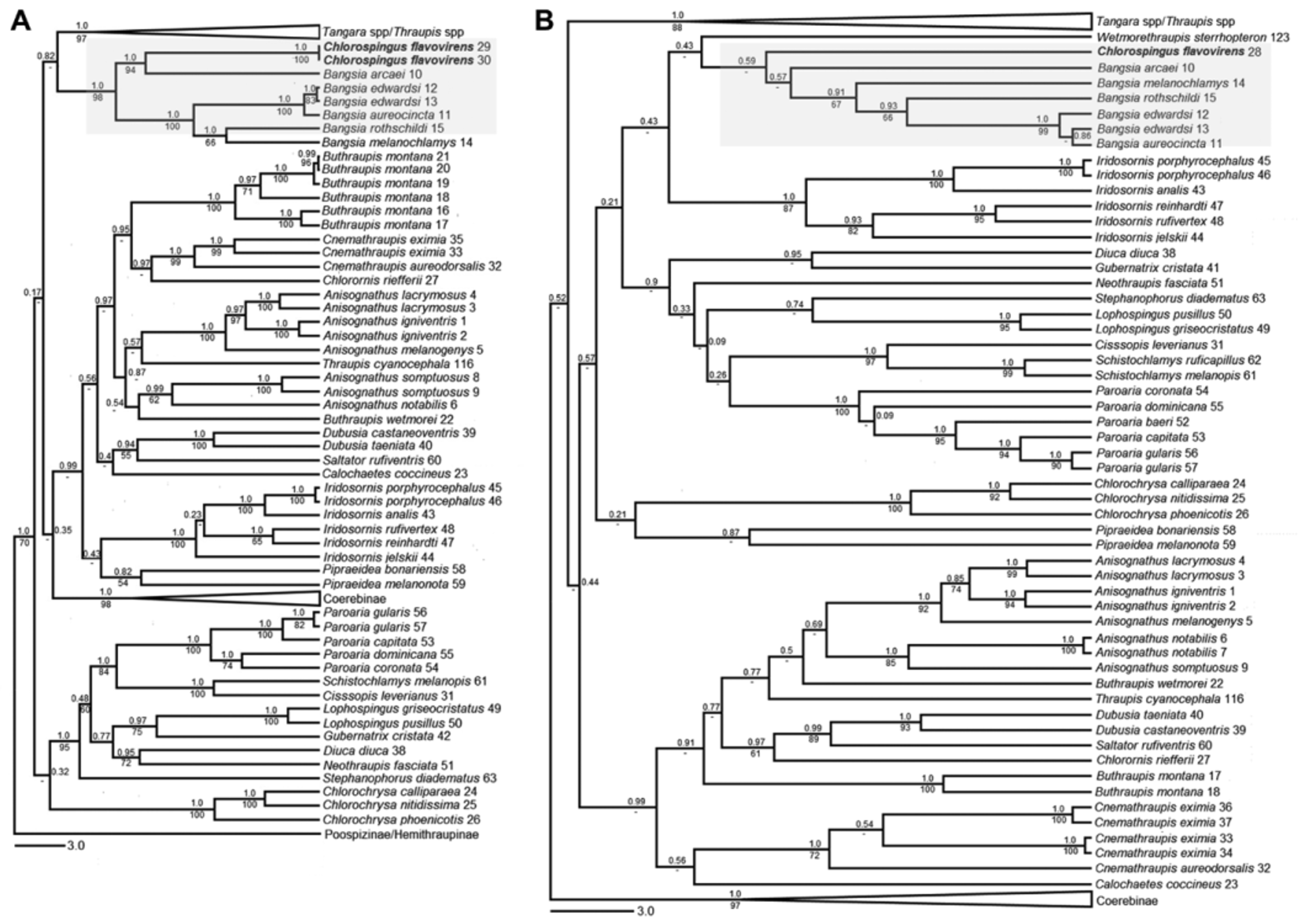
Analyses based on all three mitochondrial genes suggested that the Yellow-green Bush-tanager is not closely related to species of Chlorospingus or to any other emberizid, but instead, is deeply nested within the Thraupidae . For the ATPase 6 & 8 genes ( Fig. 1 View FIGURE 1 ), all Chlorospingus species formed a well-supported clade (posterior probability = 0.96; ML bootstrap = 50%; supports hereafter in this order) within the Emberizidae (or more specifically the Passerellidae , following the taxonomy of Barker et al. 2013), whereas C. flavovirens was included in a distantly related but weakly supported clade containing tanagers and cardinals. Within this clade, however, the species was included in a strongly supported clade (1.0; 64%) containing typical Thraupinae taxa (e.g. Buthraupis , Tangara ). ND2 analyses clearly showed C. flavovirens within the core tanager clade (1.0; 70%; Fig. 2 View FIGURE 2 A). Morever, sequences of this gene strongly suggested the species is sister to Bangsia arcaei Sclater & Salvin (1869) (1.0; 94%); the clade formed by C. flavovirens and B. arcaei was sister to a clade including the remaining species of Bangsia Penard (1.0; 98%). Relationships inferred based on the cyt b gene were not as strongly supported, especially within the core tanagers ( Fig. 2 View FIGURE 2 B). Although support was not strong in analyses based on this gene, C. flavovirens was also found to be a member of the core tanagers; in contrast to the ND2 topology, it was not recovered as sister to B. arcaei , but rather as the sister group of Bangsia , albeit with weak support. The concatenated analyses resulted in topologies and support values similar to those obtained in the ND2 analyses ( Fig. 3 View FIGURE 3 ): C. flavovirens was nested within the Bangsia clade (0.99; 81%) as the sister taxon to B. arcaei (1.0; 93%).
We used estimates of marginal likelihoods based on the concatenated data set to contrast the null hypothesis where C. flavovirens is sister to B. arcaei to the alternative hypothesis where C. flavovirens is sister to a clade formed by all species of Bangsia using the Bayes factor. Estimates of the marginal likelihood were -82045.67 for the null hypothesis and -82519.12 for the alternative hypothesis, resulting in an absolute difference (i.e. Bayes factor) of 473.45. Because Bayes factors larger than 150 should be interpreted as very strong evidence in favor of the better model ( Kass & Raftery 1995; Bergsten et al. 2013), this analysis strongly favors the hypothesis that, given our mtDNA data, C. flavovirens is sister to B. arcaei .
Chlorospingus flavovirens was originally described as Buarremon flavovirens ( Lawrence, 1867) . It was later transferred to Chlorospingus Cabanis, 1851 by Sclater (1885) based on its short and strong beak, more typical of Chlorospingus species. Since then, it has been consistently considered a Chlorospingus , sharing similar plumage patterns (particularly with some species; e.g. C. tacarcunae , C. inornatus ), habitat and behavior with members of this group ( Isler & Isler 1999; Hilty 2011; see Fig. 1 View FIGURE 1 ). Our results conclusively show that C. flavovirens is actually a tanager distantly related to true Chlorospingus , for which the type species is the taxon now known as C.
flavopectus ophthalmicus ( Du Bus, 1847) . Accordingly, the species must be transferred to the subfamily Thraupinae within the Thraupidae .
Examples of morphological similarity between phylogenetically distant taxa are not rare among Emberizoids, and this is one of the reasons why classification in groups like tanagers, emberizids and cardinalids has been subject to extensive revision following molecular phylogenetic analyses (Burns et al. 2014, 2016; Klicka et al. 2014; Barker et al. 2015; Remsen et al. 2016). However, the similarity between C. flavovirens and true Cholorospingus that had thus far misled taxonomists who considered them congeneric represents a remarkable case of evolutionary convergence in plumage, morphology, and behavior between taxa belonging to different families.
In addition to being transferred to the Thraupidae , C. flavovirens should be placed in a different genus to make classification consistent with phylogeny. The genus name Buarremon Bonaparte, 1850 in which the species was originally described is not available because the type species is Embernagra torquata Lafresnaye and d’Orbigny, 1837 , (i.e., Arremon torquatus ; Cadena & Cuervo 2010), an emberizid distantly related to C. flavovirens . Because no generic name is available for C. flavovirens , based on our results we see three alternatives: (1) creating a monotypic genus for C. flavovirens , (2) placing C. flavovirens and the closely allied B. arcaei in a genus separate from the other species of Bangsia , or (2) merging C. flavovirens and Bangsia in a single genus. We favor the third option for reasons outlined below.
Creating a monotypic genus for C. flavovirens would recognize the phenotypic and evolutionary distinctiveness of this taxon. However, given our finding of its close relation with B. arcaei , which is the type species of Bangsia (Ridgway, 1893) , this would require naming an additional genus for the remaining four species now included in this genus. This would result in recognizing three genera and in changing the genus of C. flavovirens and of four species of Bangsia , a solution that would involve substantial changes to the existing classification. In addition, this alternative would imply recognizing two monotypic genera (one for C. flavovirens , one for B. arcaei ); we favor not doing so because monotypic genera convey no information about phylogenetic relationships. Of course, classifying species in monotypic genera does convey phylogenetic information in cases where it highlights deeply divergent lineages with no close relatives (e.g., Sapayoa Hartert or Donacobius Swainson among Neotropical passerines), but we argue this is not the case for C. flavovirens .
The second alternative (i.e. merging C. flavovirens and B. arcaei in Bangsia ) would prevent the naming of a new genus for the former, but again this would require naming a new genus for the remaining four species of Bangsia . Moreover, although the ND2 data and the concatenated analyses strongly show that C. flavovirens is sister to B. arcaei , this was not supported by the cyt b data. An important consideration regarding this and the former alternatives, both of which imply separating B. arcaei from other species of Bangsia , is that this has no apparent phenotypic basis given the relatively conserved plumage and morphology across these species. Although Penard (1919) noted some differences in bill shape and primary length between B. arcaei and its congeners, neither we nor Penard could draw reliable conclusions about the taxonomic validity of these characters due to the scarcity of specimens.
Finally, merging C. flavovirens and Bangsia in a single genus would avoid naming two new genera and would imply changing the name of only one species, thus maximizing taxonomic stability. In addition, this treatment would be consistent with all of our analyses, which unequivocally place C. flavovirens in a clade with species of Bangsia regardless of its position within such clade. Despite our findings showing strong support for the position of C. flavovirens as sister to Bangsia , we do not deny the possibility that C. flavovirens may be sister to Bangsia as suggested by the cyt b data; however, if this were to be confirmed with additional analyses considering other characters (i.e. nuclear DNA data), our proposal to merge C. flavovirens and Bangsia would still render a classification consistent with phylogeny. Moreover, we suggest that this treatment is more informative about the evolutionary relationships of the taxa involved despite implying the recognition of a phenotypically heterogeneous genus. Although differences in plumage coloration between C. flavovirens and species currently included in Bangsia are rather striking ( Fig. 3 View FIGURE 3 ), all these taxa are generally similar in size and proportions, so one may think of C. flavovirens as a taxon lacking complex plumage patterns and some of the colors present in relatively close relatives such as B. edwardsi ( Elliot, 1865) or B. aureocincta ( Hellmayr, 1910) . Finally, we note that C. flavovirens occupies the center of the geographic distribution of Bangsia , reflecting a shared history of diversification in northwestern South America among members of this clade (Sedano & Burns 2010).
In summary, to make classification consistent with our current understanding of phylogeny, we suggest that the taxon heretofore referred to as Chlorospingus flavovirens (Emberizidae) should be transferred to the Thraupidae and referred to as Bangsia flavovirens . To avoid confusion with members of the genus Chlorospingus , we further suggest that its English name should be changed to Yellow-green Tanager. We encourage further studies involving a broader gene sampling, together with morphological and vocal analyses to improve our understanding of the systematic relationships and evolution of phenotypic diversity in this group.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.