
Typophyllum lacinipenne Enderlein 1917
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4012.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:4DE5E609-AC90-4AA5-84D1-AA0D86B5C4DB |
|
DOI |
https://doi.org/10.5281/zenodo.6103142 |
|
persistent identifier |
https://treatment.plazi.org/id/03A987CF-0553-A906-FF45-2841FC4B276B |
|
treatment provided by |
Plazi |
|
scientific name |
Typophyllum lacinipenne Enderlein 1917 |
| status |
|
Typophyllum lacinipenne Enderlein 1917
T. undulatum Caudell 1918 syn. nov.
T. acutum Vignon 1925 syn. nov.
http://lsid.speciesfile.org/urn:lsid: Orthoptera View in CoL .speciesfile.org:TaxonName:5490
T. lacinipenne: Enderlein 1917 ( T. lacinipennis ): 19, Vignon 125a: 270 ( T. lacinipennis ), Vignon 1931 ( T. lacinipennis ): 135, Beier 1960: 365; T. undulatum: Caudell 1918: 53 ; Vignon 1925a: 269, Vignon 1930: 427, Vignon 1931: 134, Beier 1960: 365; T. acutum: Vignon 1925a: 270 , Vignon 1931: 135, Beier 1960: 365.
Second group acording to Vignon (1925a), with basally broadened hind tibiae. All three described from unique females (photos in OSF). T. lacinipenne with mostly green tegmina is from central Bolivia (Provincia Sara). T. undulatum from central Peru is brown, slightly smaller, and has the anal margin of the tegmina distally undulated. The specimen of T. acutum , also from central Peru (Pozuzo), with distally slightly undulated tegmina, is yellowish brown and was perhaps greenish in life. Since there are only marginal differences, it does not seem very reasonable to maintain three different names. Hopefully the corresponding males will be discovered soon.
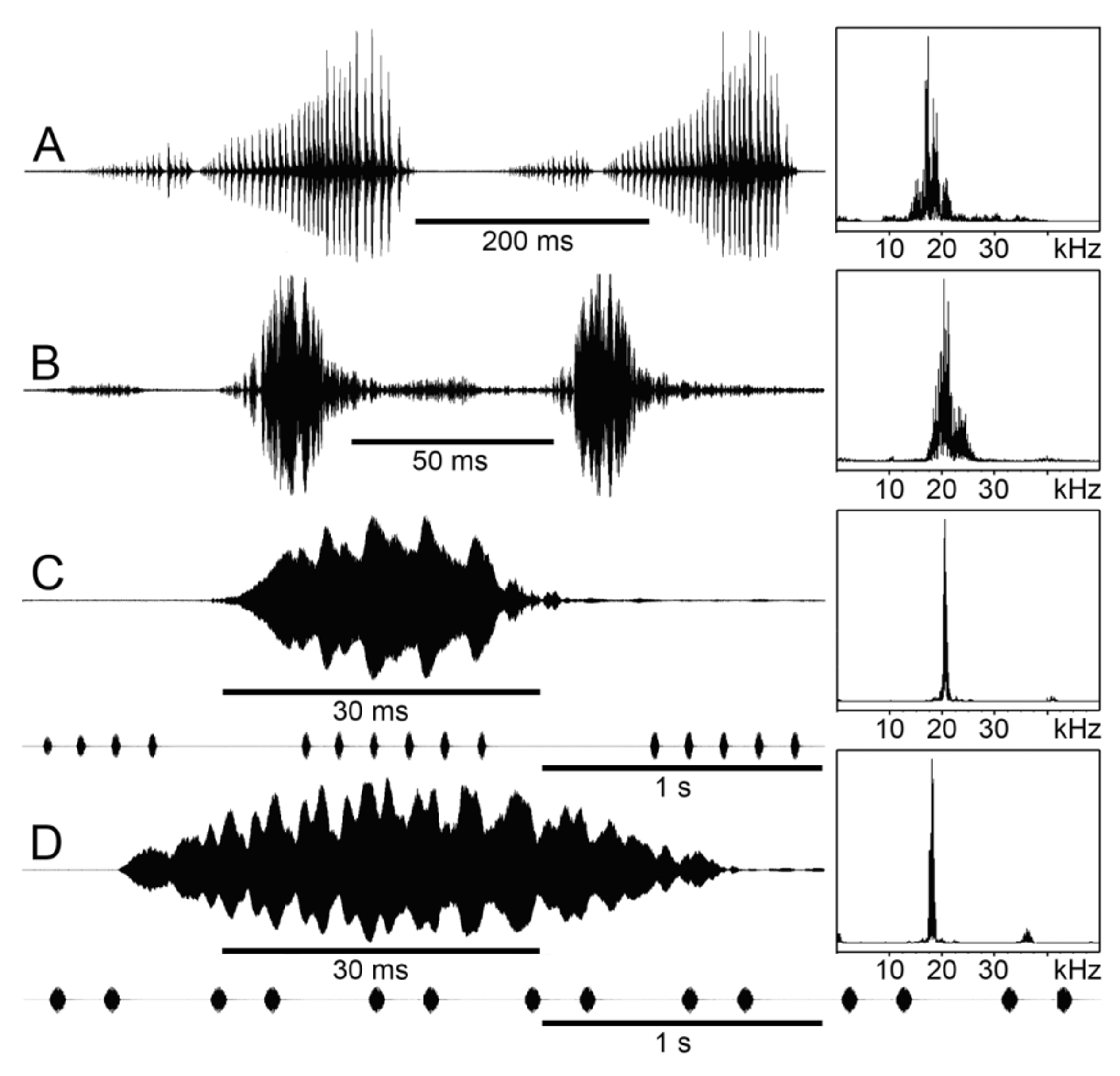
The songs. With the calling songs of four species added here, there are now documented the songs of eight or nine Typophyllum species: T. bolivari , T. mortuifolium , T. sp. nr. trapeziforme ( Morris et al. 1989, the latter could be T. morrisi sp. nov.), T. zingara (Montealegre & Morris 1999) , T. sp. ( Morris & Montealegre 2001), T. egregium , T. erosifolium , T. morrisi sp. nov. and T. onkiosternum sp. nov. (this work, Fig. 19 View FIGURE 19 ). From two other species of Pterochrozinae View in CoL the songs are known as well: Mimetica incisa ( Morris & Beier 1982) and Cycloptera arcuata ( Braun 2002, recordings in OSF). In all species the spectrum of the carrier frequencies is extraordinarily narrow, in some species composed of almost pure sine waves. In the large Cycloptera male the peak is at 10 kHz, and in M. incisa , with the male about the size of the females of small Typophyllum species (27–29 mm tegmen length, Vignon 1931), 14 kHz. In the so far studied Typophyllm males the fequency ranges from 9.5 to 23 kHz, with the smallest species calling at around 20 kHz, which is slightly above the boundary to ultrasound. In these species only very close to a calling male a faint sound is audible to the unaided ear, possibly rather a suppressed fundamental frequency than low-energy components close to the real carrier frequency. With an ultrasound detector, calling males can be heard over fairly long distances. Provided that the females can hear equally well, these maximum broadcast distances probably cover more than the range a phonotactically approaching female can move in one night through understory vegetation.
A somewhat broader carrier frequency spectrum show the partly audio-range T. egregium and the ultrasonic T. onkiosternum sp. nov. ( Fig. 19 View FIGURE 19 A,B). The distribution of T. egregium reaches up to almost 3000 m ( Fig. 20 View FIGURE 20 A), where it lives in patches of low elfin woodland ( Fig. 21 View FIGURE 21 A). T. onkiosternum sp. nov. is known from very few indivudals that were found around 2150 m. The species that produce pure tone songs, like T. erosifolium , T. bolivari and T. mortuifoium , all occur in lower elevations, where the trees are high and the understory vegetation is relatively sparse. With increasing elevation, the density of the understory vegetation increases as well, whereas the size of the trees decreases. A similar correlation with habitat structure, that open-habitat species produce songs with a broader spectrum, is reported for birds (e.g. Bertelli & Tubaru 2002). Forest bird songs tend to be more pure tone, probably due to a comparatively stable atmosphere, which permits selection to favour mechanisms to increase the sound intensity ( Morton 1975). By concentrating all the energy on a very narrow frequency band, the transmission range can be maximized ( Wiley & Richards 1978). This may be also important for the sparsely distributed forest-dwelling katydids. Broad-band songs might evolve as an adaptation for distance ranging by differential degradation of higher versus lower frequencies in dense vegetation (Montealegre & Morris 2004). This could be particularly important for females of T. egregium in the upper range of the species, where its habitat comprises patches of elfin forest. These are fairly fragmented and separated by areas with low vegetation of a small bamboo species and terrestrial bromeliads, which provide little protection against the harsh weather conditions. So it could be hazardous to approach a male calling from too far away.
In males of the species that produce pure tones, the speculum on the base of the right tegmen, that serves to amplify and spread the sound, seems to be rather stiff and is merely translucent ( Fig. 7 View FIGURE 7 B, photos of T. morrisi sp. nov. and T. erosifolium in OSF). Males of T. egregium and T. onkiosternum sp. nov., whose songs have a broader spectrum, have a fully transparent speculum ( Fig. 4 View FIGURE 4 E). This was also found in the neotropical katydid genus Panacanthus View in CoL , with pure tone resonant songs as ancestral state, where in species with non-resonant songs the speculum membranes are glassy and thin (Montealegre & Morris 2004). Such a thin speculum could be more capable of vibrating in intrinsic modes as a result of successive transient tooth-scraper impacts, while the thicker and less transparent speculum of resonant singers is more suitable for a mechanism where the membrane is driven by tooth-contact rate (op. cit.). In males of T. morrisi sp. nov. and T. erosifolium the speculum membrane looks very homogenous and must oscillate uniformly and smoothly between the solid surround. The teeth on the stridulatory file of these two species are microscopically small and very densely packed, compared to the nonresonant T. egregium , where they are not so small and tooth density and number are lower. Whereas one male of T. erosifolium has almost 300 teeth on its file, a male of T. egregium has less than 150.
The curious mating behaviour. Pre-copulatory riding of the tiny male on one side of the female’s tegmina, as observed in Typophyllum egregium and T. erosifolium , is also recorded for T. trapeziforme ( Xiberras & Ducaud 2014b), T. zingara (Montealegre & Morris 1999) , T. bolivari , T. mortuifolium , T. trigonum , as well as for Roxelana crassicornis and Pterochroza ocellata ( Castner & Nickle 1995a; whereas this is doubtful for the latter species, where the male reaches almost the same size as the female, and which apparently shows a different behaviour— P. Ducaud pers. comm.). In T. egregium the riding of the male on the female’s tegmina preceded mating, and the two always separated after having accomplished the protracted transfer of the spermatophore. Likewise proceeds T. trapeziforme , where the male also rides several days and always leaves the female after mating (P. Ducaud pers. comm.). However, Castner & Nickle (who unfortunately neither assign the behaviour to a certain species nor provide details of their nine field and more than 100 cage observations) report the male remounting the female after mating and resume riding for several hours to several days. This seems to conflict most of the speculations about the adaptiveness of this peculiar behaviour already discussed by the authors themselves (op. cit.).
One possible explanation for the pre-copulatory riding might be associated with the low probability for males and females to meet, due to the low population density, a widely acknowledged feature of arthropods in tropical forests (e.g. Elton 1973, Novotny & Basset 2000). Therefore, one or both partners might for some economical reason not always be disposed to mate immediately. But since the male does not feed while atop the female, he could only reallocate internal resources to produce a spermatophore. The female should not have any problems with receptivity, since adult female tettigoniids have a spermatheca to store sperm until use.
Another potential explanation discussed by Castner & Nickle (1995a), mate-guarding by the male, can probably also be excluded, as the peaceful contiguous riding and subsequent mating of three males with the same female in caged T. egregium showed. Furthermore, it does not seem to be necessary because of rarity in the natural habitat—‘chance encounters’ of male and female (op.cit.) are highly unlikely and the males have to rely on their calling song to bring about a rendezvous. However, if the male should actually resume the piggyback position after mating (op.cit.), it could be adaptive for the pair to stay together. It would save the female future toilsome and possibly dangerous phonotactic approaches, and the male the calling, which might attract predators or parasitoids. But if the probability for the male to attract another female before his previous mate uses up all his sperm is higher than to attract the interest of an eavesdropping bat, he should desert her (or perhaps resume calling atop).
According to another assumption (op.cit.) the female might evaluate the male by assessing his weight. But why should she venture a phonotactic approach at all (calling already can convey information on male quality, e.g. Gwynne 2001), and then carry him around for several days instead of retrieving on the spot a more or less nutritious spermathophore and get rid of him? An instance of a male T. egregium , who descended after five days of riding on the female without mating, rather suggests the possibility of the male assessing the female.
Possibly the piggyback behaviour of the little walking leaves is one facet of their sophisticated crypticity (see next section). Castner & Nickle (1995a) already presumed that a male sitting on the wings of a female might foil the search image of predators. But perhaps it is the female alone who obliges the male to ride on her. If the curious ensemble attracts the attention of a predator despite its camouflage, the small male ‘sitting on a leaf’ is spotted first, while the female can escape. For instance in stick insects survival of females can improve considerably with dorsally coupled males ( Sivinski 1983). Probably the time the male can spend riding (up to five days in T. egregium ) is limited by starvation, unless he descends from time to time to feed.
Likewise mysterious as the reason for this behaviour remains presently the question whether the sexual size dimorphism in Typophyllum , which is extraordinarily pronounced for a katydid, might be an adaptation to it. This seems to be the case in certain stick insects with prolonged mating, were the female carries around the smaller male. In those species the sexual size difference is especially distinctive ( Sivinski 1978, 1983).
Why are the little walking leaves so leaf-like? Already early orthopterists were intrigued by the perfection of mimetism in which the Pterochrozinae View in CoL excel. While the tegmina with their venation are predestined for the imitation of a leaf, as many other katydid species apparently use as simple camouflage, the little walking leaves additionally exhibit various details that mimic a real separate leaf in various stages of decay. These include tiny transparent windows, brownish patches and margins in otherwise green individuals, undulated or excised margins, light greyish spots, and minute whitish or dark tubercles—all even through a stereomicroscope looking like real holes caused by small herbivorous insects, necrotic spots, lichens, or fungi. In the larger females the dorsal edge of the tegmina is sometimes slightly twisted, so that in dorsal view the midline of the tegmina is s-shaped, just like a withered leaf. Brown body parts sometimes are partly green and “mossy”. Nymphs, that cannot mimic a leaf due to the undeveloped tegmina, frequently have light green and ramified processes on the abdominal tergites, that look like real moss ( Figs. 9 View FIGURE 9 B, 11C). The antenna tips are often curled ( Figs. 11 View FIGURE 11 D, 15G, 16), reminiscent of a tiny vine, as in other camouflaged katydids, like the little lichen dragon Lichenodraculus matti ( Phaneropterinae View in CoL , Dysoniini View in CoL ), that was also found in the investigation area ( Braun 2011).
Back in 1894, the eminent orthopterist Brunner von Wattenwyl considered these numerous details as exuberance and an effort far in excess of the necessary, which could not be attributed exclusively to natural selection. Vignon (1925a) concurred with this view and thought the luxury of accessories to be rather contrary to real mimetism. Still in his fine and beautifully illustrated monograph (1931) he came to the conclusion that the detailed resemblance to decayed leaves, or leaves apparently mined or eaten by caterpillars, was useless, his reason being that other species with the much simpler likeness to uninjured leaves are able to hold their own in the struggle with greater success, as shown by their comparative abundance. He considered the details as a decoration unnecessary in the life of the insects, a multiform comedy, mimetism and aesthetics being two aspects of a sole mystery. In contrast Karny (1914) already assumed that to deceive sharp-eyed insect-feeding birds it requires an imitation of the leaf in colour and shape up to the smallest detail. He still entertained some doubt that selection alone was sufficient for such crypticity and contemplated Lamarck’s (1809) theory of active acquisition of useful traits.
The different birds of mixed species flocks in the investigation area actually searched very meticulously bark, foliage, bromeliads and other epiphytes, including all sorts of crevices and possible hiding places of arthropod prey (pers. obs.). Since the little walking leaves furthermore come in multifarious outfits (as shown particularly in T. egregium ), it becomes even more difficult for birds as the main visual predators to develop a search image, a term introduced by Tinbergen (1960). Such polymorphism in colouration has been recorded in a variety of cryptic prey species ( Bond & Kamil 2002 and references therein) and is shown to be the potential outcome of frequency dependent selection (op. cit.), since predators have to allocate their attention economically ( Dukas 2004 and references therein). The underlying mechanisms are complicated and not fully understood ( Gray & McKinnon 2007). Through the piggyback behaviour males and females stay together during day and can perceive colour, so even some sort of “assortative” mating is thinkable. The little walking leaves held in cages did not choose their daily roosting places according to their own colour, although differently coloured substrates were available, not confirming observations for Mimetica mortuifolia (Bellwood 1988) , but consistent with other observations ( Castner & Nickle 1995a). And why should a little leaf as an object of its own have the same colour as the background anyway?
In the investigation area the little walking leaves were quite rare. This fits into an additional line of reasoning ( Hochkirch 2001): a high predation pressure combined with habitat stability might favour individuals that invest in high life expectancy (a T. erosifolium male caught as adult lived for nine more months, calling almost every night) and produce small clutches (one egg per site in unspecified Pterochrozinae View in CoL , Caster & Nickle 1995a), therefore, do not have to feed and move much. The resulting low reproduction rate would explain the low population density, which in turn complicates prey specialization by predators. Low population densities also affect evolution. Speciation might be accelerated by rarity, small distribution areas and low dispersal abilities ( Chown 1997). The underlying (and controversial, op.cit.) logic concerns gene flow, which tends to prevent speciation in abundant species with broad distribution areas ( Mayr 1963, Stanley 1979). Similar predictions make the theory of shifting balance ( Wright 1931, 1977, Eldredge 1985, Holt 1997). The potentially creative role of random genetic drift permits small populations to move between adaptive peaks, while in large populations adaptive evolution is less likely and they are in some measure evolutionary inert because of gene flow.
These merely indicated mechanisms might have contributed to the extraordinarily perfect imitation of tiny leaves, including all the microscopic accessories like factitious fungi cultures on the tegmina. If the tropical diversity is not already very old compared to temperate regions, then the speciation rate must be especially high (e.g. Chown & Gaston 2000). Additionally the predation pressure in the tropics is thought to be higher than in temperate ecosystems ( Hawkins et al. 1997 and references therein), probably again as a result of rarity of potential prey species.
Where did the walking leaves originate in the tree of life? A recent molecular phylogeny of Tettigoniidae View in CoL includes a remarkable result: a clade consisting of Mimetica tuberata from Costa Rica and Typophyllum sp. from Peru is found to be a sister group to all other katydids ( Mugleston et al. 2013). Traditionally the group was mostly treated as tribe Pterochrozini View in CoL within Pseudophyllinae View in CoL (e.g. Beier 1960), sometimes as subfamily (e.g. Gorochov 2012, there under subfamily group Pseudophyllidae ), and it was proposed to place it directly under Tettigoniidae (Braun 2015) View in CoL . One indication that the Pterochrozinae View in CoL might be unrelated to Pseudophyllinae View in CoL is the development of the non-functional stridulatory file on the right tegmen. It appears as an unsclerotized and inconspicuous vestigial copy of the functional one on the left tegmen, like in all other subfamilies investigated regarding this character, except for the true pseudophyllines, that typically have it completely reduced ( Chamorro-Rengifo et al. 2014). However, another characteristic share Typophyllum species and Pseudophyllinae View in CoL : The functional file is located all along a fairly massive cubital vein that projects basally beyond the lower tegmen surface. Another indication for an early phylogenetic origin is the pure-tone resonant stridulation, which is thought to represent an ancestral character state in katydids (Montealegre-Z. & Morris 2004, Montealegre-Z. 2009). Future studies on katydid phylogeny will hopefully substantiate how long ago these fascinating insects did branch off.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Pterochrozinae |
|
Genus |
Typophyllum lacinipenne Enderlein 1917
| Braun, Holger 2015 |
T. lacinipenne:
| Beier 1960: 365 |
| Beier 1960: 365 |
| Beier 1960: 365 |
| Vignon 1931: 134 |
| Vignon 1931: 135 |
| Vignon 1930: 427 |
| Vignon 1925: 269 |
| Vignon 1925: 270 |
| Caudell 1918: 53 |