
Hansenocaris aquila Grygier and Olesen, 2022
|
publication ID |
https://doi.org/ 10.12782/specdiv.27.301 |
|
publication LSID |
lsid:zoobank.org:pub:DE06A95A-7DC2-41BA-B94B-2E8DDA5FBE86 |
|
DOI |
https://doi.org/10.5281/zenodo.7544609 |
|
persistent identifier |
https://treatment.plazi.org/id/10952D25-5976-4F4D-B131-FE886DAC885B |
|
taxon LSID |
lsid:zoobank.org:act:10952D25-5976-4F4D-B131-FE886DAC885B |
|
treatment provided by |
Felipe |
|
scientific name |
Hansenocaris aquila Grygier and Olesen |
| status |
sp. nov. |
Hansenocaris aquila Grygier and Olesen , sp. nov. [New Japanese name: Washi-chou-kou-mushi] ( Fig. 6 View Fig )
Diagnosis. In last-stage nauplius, labrum wine-glassshaped in ventral view with rounded posteriolateral corners, median keel carrying 3–4 small spines, and robust, sharply pointed posterior spine reminiscent of eagle’s beak. Labral surface with characteristic ridge pattern described below. Cephalic shield clearly and completely reticulated. Dorsum of faciotrunk with 4 longitudinal spine rows, inner pair of rows longer than outer pair. Second antennae and mandibles devoid of feeding structures (lecithotrophic), segmentation of their exopods and endopods 6/5 and 1/1, respectively. First maxillae and dorsocaudal organ absent. Dorsocaudal spine nearly as long as trunk dorsum preceding it, armed with robust spines. Furcal spines small.
Type locality. Off pier at the University of the Ryukyus Tropical Biosphere Research Center , Sesoko Station, on Sesoko Island, Okinawa Prefecture, Japan (26°38′09.3″N 127°51′55.2″E) GoogleMaps .
Type material. Holotype: exuvium of last-stage nauplius prepared as semi-permanent glycerine jelly slide-mount, Natural History Museum of Denmark. NHMD-1174615; collected alive as young nauplius on 22 September 2005, last stage isolated from batch culture on 25 September, its empty molt retrieved on 27 September . Paratype: exuvium of laststage nauplius prepared as semi-permanent glycerine jelly slide-mount, Natural History Museum of Demark . NHMD-1174616; collected alive as young nauplius on 16–19 July 1996, final exuvium recovered on 22 July ; unclear which of 2 cyprids mounted on same slide corresponds to this nauplius . Both type specimens collected and processed by M. J. Grygier.
Etymology. The Latin name is a noun in apposition, “ aquila ” (=“eagle”), referring to the large, strongly pointed, eagle-beak-like extension of the posterior margin of the labrum. The new Japanese name combines “washi” (Japanese for “eagle,” again referring to the labrum) with an existing Japanese name for Facetotecta (“chou-kou-rui”) plus “mushi,” meaning “bug” or “worm.”
Description (holotype). A last-stage nauplius larva ( Fig. 6 View Fig ).
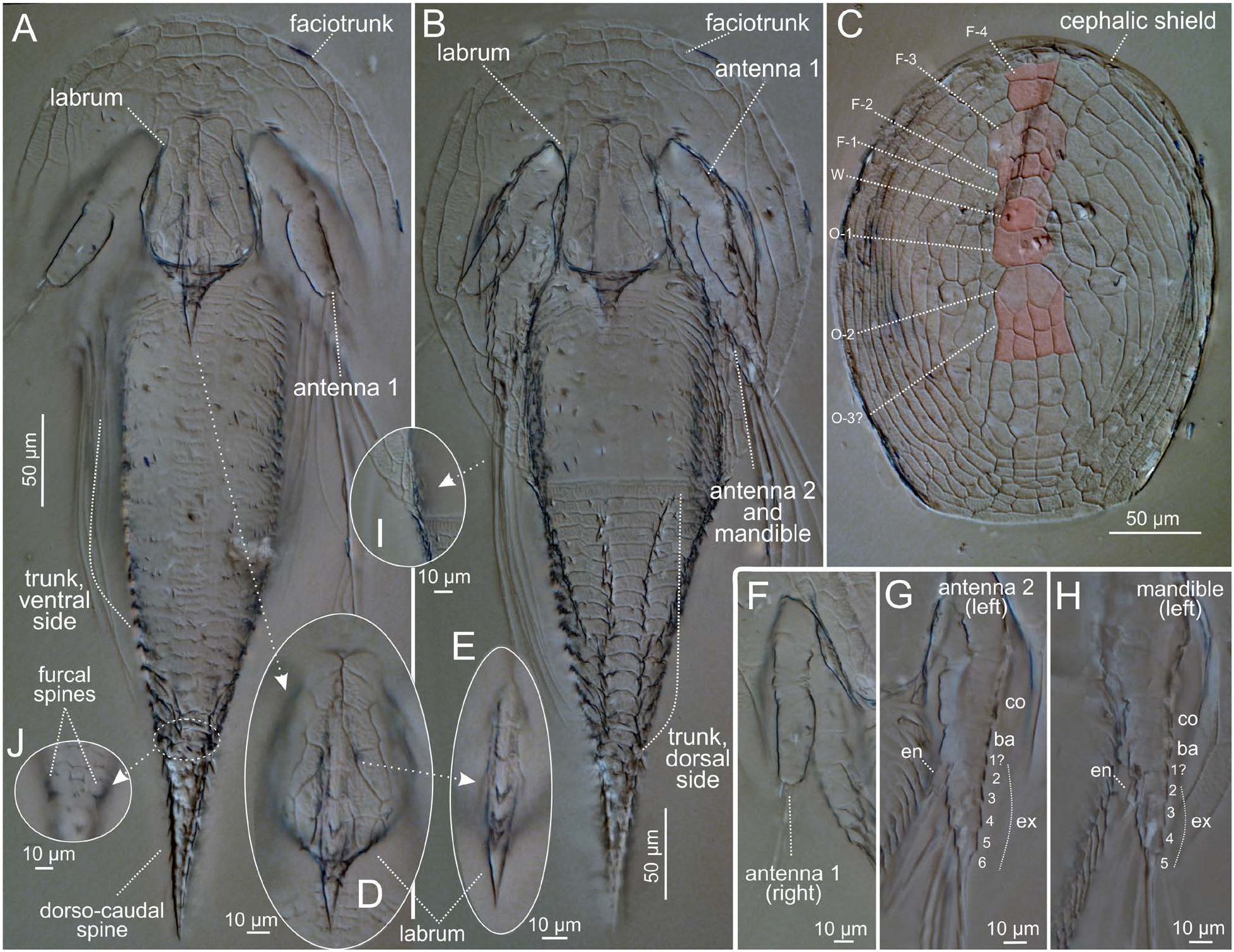
Habitus ( Fig. 6A, B View Fig ). Cephalic portion slightly oblateoval, trunk portion long and attenuate. Total length 500 µm; length and width of cephalic shield 265 and 210 µm, respectively; anterior width and post-labral length of trunk in ventral view 105 µm and 350 µm, respectively. Length (measured from furcal spines) and basal diameter (at position of furcal spines) of dorsocaudal spine 100 µm and 30 µm, respectively. No lateral view available but long axes of cephalic shield, trunk, and dorsocaudal spine apparently nearly in same plane (no significant bending).
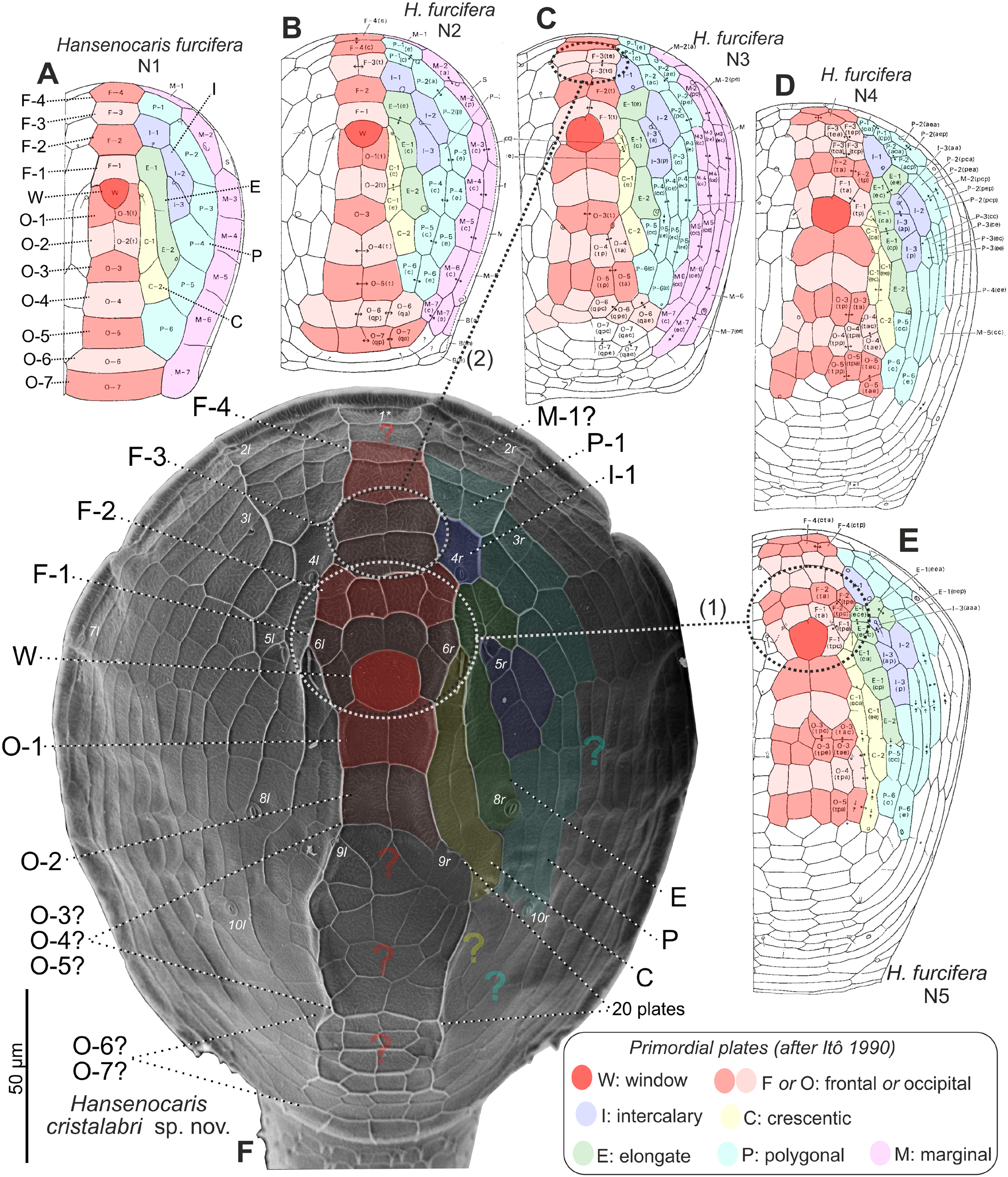
Cephalic shield ( Fig. 6C View Fig ). With dense, nearly symmetrical pattern of reticulate ridges outlining many so-called plates, or facets. Setation and pore pattern of shield not clearly visible, but posteriolateral corners lacking H. cristalabri -like pair of spine-bounded notches. Plates or facets centered on rounded-quadrangular ‘window’ (W) at about one-third length along midline, but only those near W ( Fig. 6C View Fig ) easily identifiable with those of H. cristalabri sp. nov. and H. furcifera . Namely, region of primordial ‘frontal’ plate F-1 represented by pair of small pentagonal plates flanking W and by transverse row of 4 small plates preceding these and W; region of primordial ‘frontal’ plate F-2 possibly represented by array of 6 small plates preceding F-1 region, including transverse central pair and 2 longitudinally oriented lateral pairs; and primordial ‘occipital’ plates O-1 and O-2 represented by 2 successive pairs of large plates posterior to W. Configuration of all other plates, including more anterior F-plates, more posterior O-plates, and most non-ax-ial plates, differing from those in H. cristalabri sp. nov. and H. furcifera (cf. Fig. 4E View Fig ) and, owing to lack of information about important “land-mark” pores and setae, also difficult to homologize with any other species’ primordial plates; therefore, detailed description omitted.
Labrum ( Fig. 6A, B, D, E View Fig ). Excluding its robust, sharply pointed, 35 µm long posterior extension, main portion of labrum obovate or rounded-spatulate in ventral view, 80 µm long and 60 µm wide, with median keel bearing row of 3 small but distinct and equal distal spines preceded by anoth-er minute spine. Labral surface divided by ridges into facets as follows: 2 elongated and overlapping facets along each lateral margin, with more anterior pair meeting in anterior midline; paired diagonal rows of 4 facets situated medial to these, extending from anterior midline to posteriolateral margin; 1 pair of posteriomesial facets preceding 2 small pairs flanking spine at posterior margin; and keel-bearing facet(s) along labrum’s posterior midline.
First antenna ( Fig. 6F View Fig ). Apparently 3-segmented, excluding narrow sclerites between 3 main segments. Unarmed first segment short, cylindrical (15 µm long, 20 µm in diameter). Unarmed second segment short, cylindrical (20 µm long, 17 µm in diameter). Distal segment 45 µm long, digitiform with moderate preaxial swelling of proximal 50%. This segment thickest (19 µm) at 1/3 length, with 3 apical setae: 2 long and 1 short.
Second antenna ( Fig. 6G View Fig ). Biramous with unclear proximal segmentation. Unarmed coxa about as thick as long (25 µm), unarmed basis shorter (20 µm long). Exopod 50 µm long, 6-segmented with rudimentary (perhaps not fully annular) proximal segment and further segments gradually becoming smaller distally, apparently bearing 5 setae in all. Endopod 1-segmented, cylindrical, 15 µm long and 7 µm thick, bearing 2 long apical setae.
Mandible ( Fig. 6 View Fig ). Similar to second antenna but smaller, again with unclear proximal segmentation. Coxa longer than basis (16 µm vs. 14 µm) and of slightly greater diameter (16 µm). Exopod 40 µm long, 5-segmented, apparently bearing 4 setae. Endopod 1-segmented, 10 µm long and 3–4 µm thick, bearing 2 long apical setae.
Hind part of faciotrunk ( Fig. 6A, B, J View Fig ). Trunk divisible into long, sparsely ornamented anterior part, heavily and concentrically ornamented posterior part, and heavily armed dorsocaudal spine. Anterior 15% or so of venter with short and sparse transverse ridges, followed by somewhat swollen and rounded middle region with short transverse ridges along midline and, more laterally, paired rows of bumps evidently representing future thoracopodal setae; in posterior third, concentric ridges well expressed and bearing small spinules ( Fig. 6A, B View Fig ). Trunk dorsum with 2 dorsal rows of spines along nearly whole length from first or second transverse ridge to base of dorsocaudal spine, with slight discontinuity posterior to midlength, and additional shorter dorsolateral pair of spine rows reaching only to dorsal rows’ points of discontinuity (4 spine rows in total) ( Fig. 6B View Fig ). Dorsocaudal organ (or positionally equivalent mid-dorsal trunk pore) absent. Dorsocaudal spine ( Fig. 6A, B View Fig ) armed with large subsidiary spines along entire length, except at sharply pointed tip. Pair of small (7 µm long), pointed furcal spines arising anterioventrally to base of dorsocaudal spine ( Fig. 6J View Fig ). Midventral (anal?) pore not observed.
No “ghost” of cyprid thorax (see Grygier et al. 2019) detected inside slide-mounted exuvium.
Description (Paratype). Trunk region rotated on slide relative to cephalic region, and posteriolateral part of one side of cephalic shield distorted; therefore, some of following measurements probably different in life. Total length 530 µm; cephalic shield length along midline 277 µm, maximal width 215 µm, posterior width 147 µm; dorsal trunk length 257 µm including 97 µm long dorsocaudal spine. Dorsocaudal spine similarly spiny to that of holotype. Labrum 113 µm long including posterior medial spine, maximal width 70 µm; main portion with same roundedspatulate or obovate shape as that of holotype and with row of small spines preceding robust, beak-like posterior extension, latter relatively shorter (23 µm) than that of holotype; cuticular ridge pattern of labrum resembling that of holotype. Four longitudinal rows of spines on trunk dorsum: 2 inner rows extending from about fourth transverse ridge to base of dorsocaudal spine, 2 outer rows extending from anterior margin only half this distance posteriorly. Limb setation as follows: first antenna with 1 long and 1 medium-long apical setae; second antenna with unarmed coxa and basis, 2 apical setae on 1-segmented endopod, and 5 setae on 6-segmented exopod (0-0-1-1-1-2); mandible similar but exopod 5-segmented with 1-1-1-2 setal arrangement. No cyprid “ghost” visible within exuvium, but pair of oval (19.5×5.5µm), purportedly “ghost”-related anterioventral invaginations present on trunk ( Fig. 6B View Fig ).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Facetotecta |
|
Family |
|
|
Genus |