
Phymaturus sinervoi, Scolaro, José Alejandro, Cruz, Fausto Méndez De La & Ibargüengoytía, Nora R., 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.214551 |
|
DOI |
https://doi.org/10.5281/zenodo.6167386 |
|
persistent identifier |
https://treatment.plazi.org/id/03AAB765-FF83-D119-FF42-FA652E750AA3 |
|
treatment provided by |
Plazi |
|
scientific name |
Phymaturus sinervoi |
| status |
sp. nov. |
Phymaturus sinervoi sp. nov.
( Figures 3 View FIGURE 3 and 4 View FIGURE 4 )
Type material. Holotype: MLP-R. 5660, adult male, collected in rocky outcrops (1000 m asl) of Cari Laufquen basaltic Tableland in Abi-Saad farm (41°02'12"S, 70°24'30.6"W), adjacent to Provincial Road 6, 61 km north of Ingeniero Jacobacci town, Rio Negro Province, Argentina. Collected by J.A. Scolaro, B. Sinervo, F. Méndez de la Cruz, J. Gutiérrez and N. R. Ibargüengoytía, December 14, 2010.
Paratypes: MLP-R. 5661, adult male; MLP-R. 5662, adult female; MLP-R. 5663, adult female; MLP-R. 5664, adult male; JAS-DC 1887, adult female; JAS-DC 1896, adult male; UNCo-PH 1899, adult male; UNCo-PH 1839 adult female; UNCo-PH 1894, adult female; UNCo-PH 1895, adult male; UNCo-PH 1902, adult male. The same data as detailed for the holotype.
Etymology. The species name is dedicated to honour our friend and colleague Dr. Barry Sinervo from the Department of Ecology and Evolutionary Biology, University of California ( USA), Santa Cruz; an evolutionary biologist and herpetologist who has enthusiastically and generously collaborated in many research projects in Patagonian herpetofauna for the last years.
Diagnosis. Phymaturus sinervoi is a member of the patagonicus group, distinguished from the neighbouring Phymaturus species by a peculiar colour pattern similar in both sexes ( Fig. 3 View FIGURE 3 ). The new species can be distinguished from the palluma = flagellifer group by having flat imbricate superciliaries rather than being rectangular and non-overlapping; slightly spiny and non-rugose caudal scales in verticilles (as seen among members of the palluma group). However, its subocular scale is not fragmented and it is separated from supralabials by two rows of lorilabials, as in most species of the patagonicus clade, but not seen in the majority of members of the palluma group. From the three geographically closer species, P. etheridgei , P. ceii and P. spectabilis , the new species P. sinervoi can be differentiated by its pattern of coloration ( Fig. 5 View FIGURE 5 ) as well as the morphological differences in snout-vent length, head length and width, hind or forelimb, finger and fourth toe length, axila-groin distance, finger lamellae, and in scales number, such as in supralabial, infralabial, lorilabial, scales around mental, subocular, and ventral scales (see results detailed above). P. sinervoi shows mainly a major body size and larger limbs, but minor number of ventral scales, when compared with P. c ei i and P. etheridgei . The same results are obtained when compared with P. spectabilis , but showing a minor number of subocular and lorilabial rows of scales.
Description of the holotype. A medium-sized lizard; snout-vent length (SVL) 90.4 mm; tail 100.4 mm; head length 16.0 mm; head width 16.5 mm; eye-nose distance 6.0 mm; forelimb length measured from the insertion of the limb into the body wall to the end of the claw of the fourth finger, 32.0 mm; hind limb length measured from the insertion of the limb into the body wall to the end of the claw of the fourth toe, 45.5 mm; axilla-groin distance 42.0 mm (45.5% of SVL); fourth finger length 10.5 mm; fourth toe length 14.3 mm; scales in dorsal head 23; scales around midbody 248; ventral scales between mental and precloacal pores 190; scales between rostral and frontal 17; supralabial scales 9-9; infralabial scales 8-8; subdigital lamellae on fourth finger 23; subdigital lamellae on fourth toe 27; precloacal pores 10; cephalic scales subhexagonals/pentagonals, smooth; supraorbital semicircles with large bulky scales, rounded, without azygous, incomplete posteriorly on both sides; no distinct rounded supraoculars; 10–11 imbricate and enlarged upper ciliaries; subocular scales rectangular, almost irregular but not fragmented, shorter than eye diameter, separated from supralabials by 2-2 irregular rows of lorilabials; preocular in contact with first lorilabial row; canthal separated from nasal by two scales; temporals smooth and rounded irregularly coniform, in 8–9 scales from auditory opening to the subocular; external auditory meatus enlarged, subellipsoidal longitudinally, with 4–5 very protruding or conically enlarged scales on its anterior border; diminute granular scales on posterior border; rostral undivided, wider twice than higher, separated by one row of medium scales from nasals; nasal large and surrounded by nine small scales; nasals separated by four small irregular scales; nostril rounded and large, over the centre of nasal scale; parietals irregular and smooth with evident subhexagonal interparietal, surrounded by seven scales; nuchals strongly conical organized in 15–16 irregular rows; post-auricular folds very developed with smooth conical scales; mental subpentagonal shorter than width, but higher than rostral, in contact with four irregular rectangular scales; two rows of 5–6 bilateral postmentals decreasing behind; dorsal scales smooth, conics, small and juxtaposed; mid-dorsal scales slightly rounded and smooth, decreasingly smaller and evidently conical toward the flanks; ventro-laterals and ventrals larger than dorsals, almost pentagonal, imbricate and smooth; two gular folds with rounded, small scales; 79 gulars between auditory meatus; caudal scales quadrangular and regularly imbricate in verticiles, proximally large, conical and smooth on dorsum, or slightly keeled, distally more rectangular and strongly keeled; scales on forelimbs sub-triangular and smooth in the upper side, granular, rounded and subconical in the under side; scales in hind limbs strongly conical and strongly keeled in the dorsum but larger subpentagonal, imbricate and flat in the under side; in the femoral region, small granular, rounded and smooth scales in the lower side; infracarpals and infratarsals with round margins, becoming more keeled to the base of fingers and toes. Subdigital lamellae of fingers keeled; fourth toe and finger claws very developed, almost 3–4 mm of long. Ten orange-yellowish precloacal glands on the scales of the cloacal region in one divided row.
Coloration. Colour pattern is similar in both sexes. The general pattern is characterized by irregularly spread pale-creamy or grey spots on a darker brownish dorsal background. These multiple clear spots are conspicuous between the dorsal area of the head, flanks, dorsal limbs, until the base of the tail, where they disappear gradually on the dorsal tail background. On the neck and fore-back, several black stripes result in a partially greyish background. The dorsal tail shows variations from entirely brown to several alternate rings (20–23) of soft light brown of one scale among dark brown rings of two scales.
On the ventral surface, the background colour varies from pale-grey, yellow, brick-red to intense orange. Colour pictures of males and females are shown in Figure 4 View FIGURE 4 .
Morphological variation. The sample analyzed comprised 14 adult males and 12 adult females (for means and SD see Table 1). Analyses show slight size differences between the sexes, females being slightly larger in SVL (79.8–103.9) than males (84.8–98.7; Table 1). Axilla-groin distance larger in females (P <0.001, Student t test: range 39.6–59.3 mm; mean = 51.4 mm, SD = 5.6, representing 53.0–58.2% of SVL); in males ranged = 42.0– 48.1 mm (mean = 45.2 mm; SD = 1.8, representing 46.5–54.6% of SVL). In both sexes, head length ranged 15.6–18.9 mm, representing 16.7–20.8% of SVL. Head width ranged 14.6–17.6 mm, with significant differences between sexes, having the males larger head width (P <0.002, t Student test). Eye-nostril distance ranged 4.5–6.5 mm. Tail length ranged 100–108.0 mm, representing 1.11–1.20 times of SVL. Forelimb length ranged 29.7–36.1 mm. Mean of hindlimb length in males was 47.8 mm, but in females 47.2 mm. Scales around midbody ranged 213–248 in both sexes combined. Dorsal head scales ranged 18–24. Ventrals ranged 157–197. Precloacal glands observed only in males, and ranged 6–13. Subocular scales not fragmented. Two rows of lorilabials between suboculars and supralabials. Scales surrounding interparietal 6–9. Scales contacting mental 4–6. Scales between rostral-interparietal 14–18. Fourth finger subdigital lamellae number 22–28. Fourth toe subdigital lamellae number 26–30. Supralabials more abundant in males (ranged = 8–10) than in females (ranged = 7–9: P <0.05, t Student test).
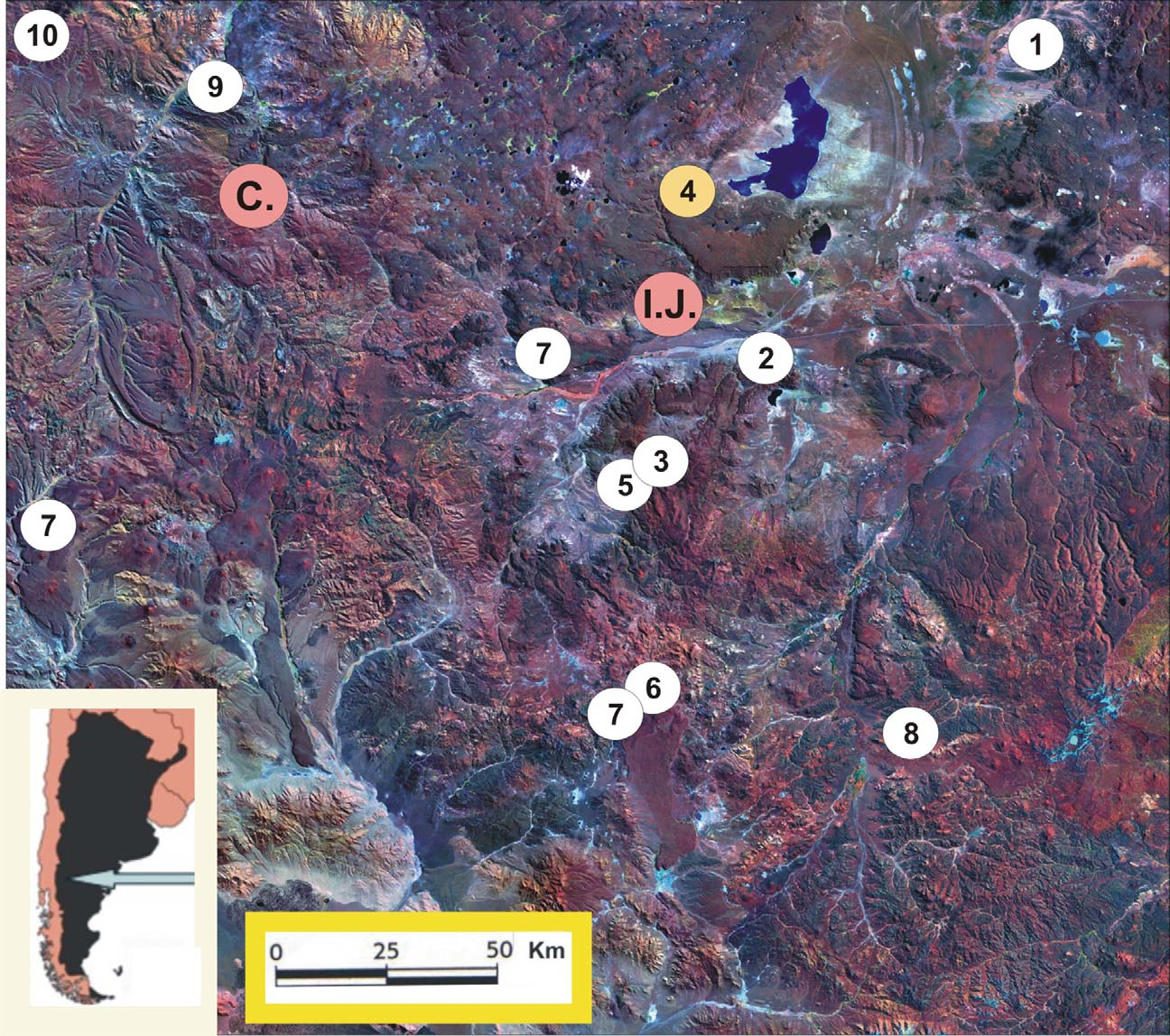
Geographic distribution. Phymaturus sinervoi has been found in an isolated biotope of the tableland of the type locality, which limits with the paleo-endorreic Cari Laufquen Grande Lagoon in the north-western side, and with sandy steppe in the south-western side, which constitute a natural barrier for the species ( Fig. 1 View FIGURE 1 ). The new species, according to the saxatil character of the species, inhabits the outcrops on the foothills of the Cari Laufquen Plateu, which shows a slight increase in height towards the south, as the result of profuse basaltic lava flows of vulcanism in the Tertiary. Plateaus are observed in staggering, product pyroclastic intercalations result of the basaltic pulses (Training Collon Cura) ( Coira, 1979). Considering the high sampling effort done for the present study we can conclude the type locality is isolated geographically and ecologically from other Phymaturus species. The type locality limits North-East with the lagoon and is surrounded by steppe shrubs typical of the low Austral Monte, where Phymaturus doesn´t advance. We also explored northward along the number 6 provincial route by sampling outcrops dispersed in the middle sandy steppe and the South-East side were only few outcrops were found, without findings of P. sinervoi . Toward the South, there is the Jacobacci town and outcrops, several times surveyed, are inhabited by of P. spectabilis , P. agilis , P. excelsus , and P. spurcus , all in varying degrees of volcanic plateaus isolation between them (see Scolaro & Ibargüengoytía 2007, Scolaro et al. 2008).
Natural history. The biotope of P. sinervoi is located inside the arid district of the Monte Austral, a steppe showing open ground, with gravel and effusive rocks. The dominant landscape is the barren steppe, with shrubby, low herbaceous coverage, with bare soil percentages above 50%. The dominant vegetation is composed by cushion bushes and sparse large clumps, the floristic physiognomy dominion are low shrubby steppes (with Nassauvia glomerulosa “colapiche”, Chuquiraga oposittifolia “quilimbay”, Mulinum spinosum “neneo”, Senecio filaginoides “charcao”, Stillingia patagonica “mata loca”, Junellia spp ., Amsinkia calycina “ortiga”, Acaena splendens “abrojo”, Perezia spp ., Ephedra spp . “solupe”, Fabiana patagonica , Grindelia coronensis “melosa”, Austrocactus patagonicus “cactus”), and mean shrubby-grass steppes (with Prosopis denudans “algarrobillo patagónico”, Lycium chilense “yaoyín”, Lycium gillesianum, Adesmia patagonica , A. volckmanni “mamuel choique”, Schinus johnstonii “molle”, and bund grasses ( Stipa papposa “coirón”, Poa ligularis “coirón poa”, Bromus catharticus ) ( Cabrera 1971; León et al. 1988).
Phymaturus sinervoi selects rocky microhabitats that occur in an isolated basaltic plateau, at elevations over 1000 m. The geological history of this geographical zone has originated several similar altitudinal plateaus, where this and other lizard species tend to live relatively isolated ( Scolaro 2006). Other reptile species coexisting with P. sinervoi are the iguanids Liolaemus elongatus , L. bibronii , L. rothi , L. inacayali , L. casamiquelai , L. ceii, Diplolaemus sexcinctus and the gekkonid Homonota darwinii . The colubrid snakes Philodryas patagoniensis and Philodryas trilineata , and the viperid Bothrops ammodytoides are also common in the same locality, representing potential predators.
Our field and lab observations reveal that P. sinervoi is viviparous, as observed in all the other species of the genus. In captivity, two females gave birth to one and two fully developed offspring respectively early in February 2011. In the field, this species is often found eating plants, as also observed in other members of the genus, i.e. Lycium chilense and Lycium gillesianum to feed on fresh fruits and flowers, and over the herbaceous species Amsinkia calycima (Scolaro et al. 2008). In lab, specimens were feed with mealworms ( Tenebrio molitor ).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.