
Unixenus intragramineus, Huynh, 2018
|
publication ID |
https://doi.org/10.11646/zootaxa.4402.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:CEA5700E-9F94-42A8-9067-BD8F4D301B9D |
|
DOI |
https://doi.org/10.5281/zenodo.5995384 |
|
persistent identifier |
https://treatment.plazi.org/id/03AB2952-FFEC-5E5B-0FBD-FB89FC930C1C |
|
treatment provided by |
Plazi |
|
scientific name |
Unixenus intragramineus |
| status |
sp. nov. |
Unixenus intragramineus sp. n.
( Fig. 2 View FIGURE 2 )
Holotype. Male, Tranh River shore (Cua Lap hamlet, Duong To commune, Phu Quoc Island , Vietnam), 10°10'22.82"N, 103°58'08.14"E; elevation 5 m; the specimen was collected by the first author from within the stem of a creeping grass ( Poaceae ) ( Fig. 3 View FIGURE 3 ) growing within the intertidal zone on the beach; on 1 st July, 2015. Queensland Museum accession number: QMS 108520 (deposited in Queensland Museum, Brisbane, Australia). GoogleMaps
Paratypes. 4 males and 6 females were collected in the same place and on the same date as the holotype. Queensland Museum accession numbers of 10 paratypes: QMS 108521–108530 (deposited in Queensland Museum, Brisbane, Australia) .
Etymology. The species is named intragramineus, noun in apposition, as it was found living in the stem of a creeping grass growing in the intertidal zone.
Diagnosis. U. intragramineus sp. n. has the general characteristics of Unixenus and it shares most characters with U. mjoebergi . Chaetotaxy: one seta present on prefemur and two setae on femur, a single seta on tibia and tarsus 2. The claw of U. intragramineus sp. n. is different from that of U. mjoebergi in being slender, with a posterior lateral process longer by half the length of the claw. A small anterior lateral process is present, anterior setiform process longer than the claw and with enlarged base.
Description. Measurements: Holotype male body length 1.7 mm, males ( paratypes) ranging from 1.7–2.0 mm ( n = 10) and females ( n = 10) from 2.2–2.5 mm. Caudal bundle of male slightly narrower in width and longer ( 0.6 mm) compared to female caudal bundle, the latter wider and shorter ( 0.5 mm).
Colouration: Head dark brown in eye area connected to 2 brown transverse bands in vertex area. Body milky white in colour, contrasting with light brown pleural trichomes and darker colour in caudal bundle; dark brown marks on latero-posterior rosette trichomes forming a dark band along each side of body laterally; last tergite darkest in colour. Ventral side of body white in colour ( Fig. 2 View FIGURE 2 ).
Head: 8 ommatidia on each side: 4 dorsal, 4 lateral (1 anterior, 2 medial and 1 posterior). Vertex with 2 posterior trichome groups and a large gap between them, each group with 2 rows. Anterior, oblique row with trichome sockets of similar size. Posterior row with slightly smaller trichome sockets. A narrow space between anterior and posterior rows ( Figs 4A View FIGURE 4 and 5A View FIGURE 5 ). Holotype with 13+13 trichome sockets in anterior row and 7+7 trichome sockets in posterior one. Paratypes with variations common in this species regardless of sex, from 10–16 (anterior rows) and 5–8 (posterior rows). Trichobothria: trichobothrium a (posterior position) and trichobothrium b (lateral position) typically thin sensory hairs with narrow cylindrical funicles; trichobothrium c (anterior position) similar in structure to trichobothria a and b, but its funicle slightly enlarged distally. All these trichobothria ( a, b and c) equal in socket size and forming an isosceles triangle with equal distance between ab and bc ( Figs 4E View FIGURE 4 and 5B View FIGURE 5 ).
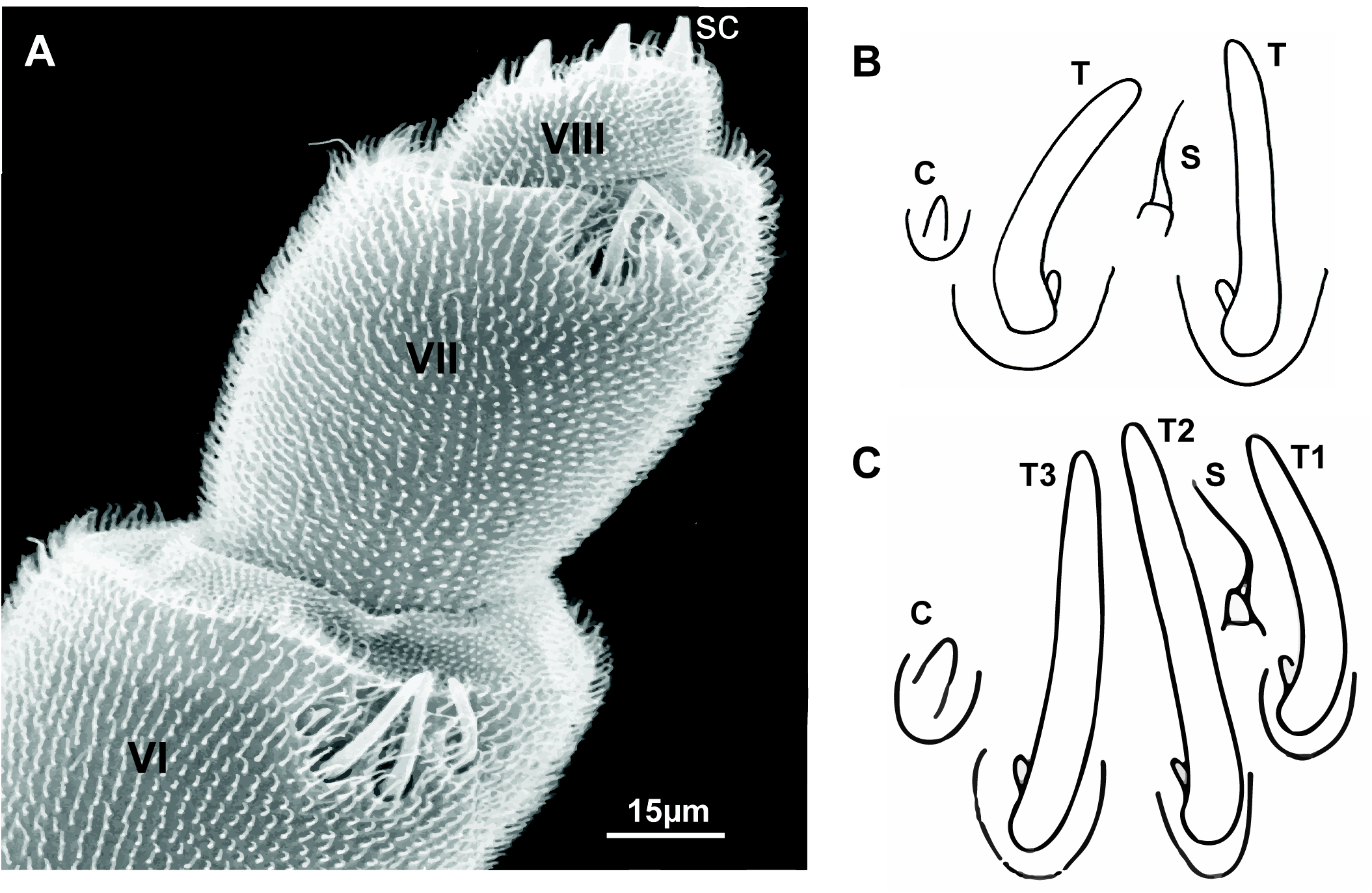
Antennae: 8 articles and 4 sensory cones ( Fig. 6A View FIGURE 6 ), characteristics typical of Polyxenidae . Antennal article VI with 3 thick bacilliform sensilla (T) of different lengths: a short sensillum anteriorly (T1), the longest intermediate sensillum (T2), and a sensillum of medium length (T3). A setiform sensillum (s) present between T1 and T2, and a conical sensillum posteriorly (c) ( Fig. 6C View FIGURE 6 ). Antennal article VII with 2 thick bacilliform sensilla (T), anterior one being slightly longer than the one located posteriorly, with one setiform sensillum (s) between them and a conical sensillum (c) located in posterior position ( Fig. 6B View FIGURE 6 ). This pattern of sensilla on the antennal article VII is commonly seen in all Unixenus species.
Clypeolabrum: Holotype with 10 setae at posterior margin, these setae being half the width of labrum. Setae in paratypes ranging from 10–15 in both sexes. Labral surface with spherical papillae with pointed ends, reduced in size posteriorly. Anterior margin of labrum with 4+4 lamellae on each side of median cleft ( Figs 4G View FIGURE 4 and 5C, D View FIGURE 5 ).
Lateral palp of gnathochilarium: Lateral palp 2.5 times as long as medial palp. 13 conical sensilla on lateral palp, medial palp with 22 slender conical sensilla observed in all paratypes and holotype ( Fig. 4F View FIGURE 4 ).
Trunk: Body with 10 segments, 9 pleural projections, excluding both telson and caudal bundle; 13 pairs of legs. Collum (tergite 1, smallest) with trichome sockets arranged in 2 oval shapes laterally, opposite each other connected by a posterior row of trichome sockets forming a line with a gap in middle. Collum, a tergite only with lateral protuberances, bearing a small number of trichome sockets. In holotype, collum with 46 (L) or 48 (R) trichome sockets, lateral protuberances with 5 trichome sockets on each side ( Fig. 4B View FIGURE 4 ). Trichome sockets on collum varying in paratypes and ranging from 42–60, trichome sockets ranging from 4–7 on lateral protuberances. All other tergites, from 2 to 10, each with a pair of pleural projections located anterolaterally. Tergal trichome socket arrangements from tergites 2 to 9 typically with 4 undefined rows of tergal trichomes arranged in two lateroposterior clusters, either side of midline. Anterior row often uneven, intermediate rows rarely in defined rows and posterior row of trichomes continuous and evenly distributed, forming a line with a medial gap. Tergite 10 exceptional in trichome sockets being smaller and denser. Almost no space between lateral rosette trichome sockets and those in posterior row. Trichome sockets of tergite 2 in holotype numbered 52 (L) and 54 (R) ( Fig. 4C View FIGURE 4 ), tergite 10 with 50 (L) and 49 (R) ( Fig. 4D View FIGURE 4 ). In contrast, trichome sockets on tergite 2 in paratypes ranging from 46–67, whereas tergite 10 containing between 53–66 trichome sockets.
Legs: Leg segments are named following Manton (1956). Legs 1 and 2 without trochanter, leg 1 lacking tarsus 1 as well. Chaetotaxy: coxa 1: 2 setae, coxa 2: 2 setae, coxae 3–13: 3–4 setae; prefemur, tibia and tarsus 2 with 1 seta; femur with 2 setae ( Fig. 7A View FIGURE 7 ). Setae on coxa and prefemur, and at distal edge of femur: bi-articulated with longitudinal ridges on basal funicle, each ridge extending distally into a long and thin spine, with the spines surrounding a base of flagellum ( Fig. 7B View FIGURE 7 ); seta of mid femur similar, but smaller ( Fig. 7C View FIGURE 7 ), tibia and tarsus 2 each with a setiform seta ( Figs 7D and 7E View FIGURE 7 ). Posterior edge of last sternite with 2–4 setae similar to those present on coxa, number of these setae varying; two present in holotype and 2–4 in paratypes. Sex organs in male: A pair of penes present on coxae 2 and 2 pairs of coxal glands located on coxal plates of legs 8 and 9.
Telotarsus-Claw: A slender structure bearing a posterior process, the latter longer by half the length of claw. Both a small anterior lateral process and a lamella process present, anterior setiform process enlarged at base and longer than the claw ( Fig. 7F View FIGURE 7 ).
Telson: Dorsal ornamental trichome sockets arranged symmetrically with 4 sockets of trichomes a in holotype; paratypes with 5–7 sockets of trichomes a. A single trichome b and 3 large protruding base sockets of trichome c: c 1, c 2 and c 3, forming a triangular shape, on each side of telson. Circular indentation d apparent near exterior side of trichomes c ( Fig. 7G View FIGURE 7 ).
Caudal bundles: In males, caudal bundle comprised of a group of caudal trichome sockets forming a single structure of uniform size; this structure split ventrally with trichome socket-free tissue and extending towards the centre with a small gap dorsally. Three rows of the largest sockets of barbate trichomes forming slightly uneven lateral rows, the latter extending towards the centre of caudal structure ( Fig. 5E View FIGURE 5 ). In females, caudal bundle different from those in males, with two obvious distinguishing features: caudal bundle structure, the main dorsal structure similar to that of male, and 2 laterosternal bundle structures with smaller sockets of nest trichomes. These smaller sockets located internally and surrounded by 2 rows of caudal trichome sockets externally. Trichome socket-free tissue present ventrally, extending with a small gap and connecting to central tissue where some barbate trichome sockets present. No separation between caudal and nest trichome sockets, all fused into a unified structure of a single caudal bundle structure ( Fig. 5F View FIGURE 5 ). Arrangement of caudal bundle of U. intragramineus sp. n. similar in structure to that observed in U. broelemanni ( Condé and Nguyen Duy-Jacquemin, 1992) , classified as type II.
Remarks. Unixenus species show very similar taxonomically important morphological characteristics: the number of sensilla on antennal article VI, the structure of the gnathochilarium and the chaetotaxy. Caudal bundle type II is common to the genus Unixenus , as well as in other genera in the subfamily Monographinae ( Condé and Nguyen Duy-Jacquemin 2008) . If only these characteristics are used for identification, it can be difficult to confidently identify Unixenus specimens to the species level. The telotarsus has proved to be a useful additional structure that can help distinguish between species in this genus. Unixenus intragramineus sp. n. has characteristics that are very similar to those of U. mjoebergi ; both species have the same chaetotaxy which makes separating these species difficult. However, the structure of the telotarsus in these two species differs. U. mjoebergi has a robust claw, while the claw of U. intragramineus sp. n. is a slender structure. U. intragramineus sp. n. is the only penicillate millipede found living within a grass stem, which is a highly specialized habitat that provides a food source and a refuge from the intermittent tidal inundation.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Penicillata |
|
Order |
|
|
SuperFamily |
Polyxenoidea |
|
Family |
|
|
Genus |