
Rhinolophus namuli sp. nov. Curran, Kopp, Ruedi, and Bayliss, 2022
|
publication ID |
https://doi.org/ 10.3161/15081109ACC2022.24.1.002 |
|
DOI |
https://doi.org/10.5281/zenodo.7738085 |
|
persistent identifier |
https://treatment.plazi.org/id/03AB87F9-FFB1-9675-7FAC-9D8A87CAF833 |
|
treatment provided by |
Juliana |
|
scientific name |
Rhinolophus namuli sp. nov. Curran, Kopp, Ruedi, and Bayliss, 2022 |
| status |
sp. nov. |
Rhinolophus namuli sp. nov. Curran, Kopp, Ruedi, and Bayliss, 2022
Common name: Namuli Horseshoe Bat
2009 Rhinolophus sp. — Timberlake et al. (2008): Mount Namuli, Mozambique: biodiversity and conservation. Report produced under the Darwin Initiative Award 15/036. Pp. 60 and 110.
2010 Rhinolophus cf. maendeleo View in CoL — Monadjem et al. (2010 b): Act. Chiropterol., 12: 371–391 — partim: Mount Namuli.
2010 Rhinolophus maendeleo View in CoL — Monadjem et al. (2010 b): Bats of Southern and Central Africa: a biogeographic and taxonomic synthesis. Wits University Press, Johannesburg: 220–223, 560 — partim: Mount Namuli.
2019 Rhinolophus cf. denti View in CoL / simulator View in CoL — Demos et al. (2019). Molecular phylogenetics of the African horseshoe bats ( Chiroptera View in CoL : Rhinolophidae View in CoL ): expanded geographic and taxonomic sampling of the Afrotropics. BMC Evolutionary Biology, 19: 166.
Holotype.— Male adult (alcohol preserved, skull, baculum preserved in glycerol), MHNG 1971.068 (field collection number MZ 120). Mozambique, Zambezia Province, Mount Namuli, Ukalini saddle forest, below mountain peak, 15.369250°S, 37.061361°E ca. 1650 ma.s.l., 21 November 2008. Captured in 2-tiered harp trap set over a small stream leading to large clearing in montane forest, at ca. 18h00. Reference sequences of the mitochondrial CYT-B and COI genes issued from the holotype are deposited in the GenBank under accession numbers MZ936306 View Materials and MZ936310 View Materials , respectively GoogleMaps .
Paratypes.— Male adult (alcohol preserved, skull, baculum preserved in glycerol), MHNG 1971.069 (field collection number MZ257). GoogleMaps Mozambique, Zambezia Province, Mount Namuli, Ukalini saddle forest, below mountain peak, 15.369250°S, 37.061361°E, ca. 1650 ma.s.l., 21 November 2008. GoogleMaps Captured in 2-tiered harp trap over small stream leading to large clearing in montane forest, ca. 18h00. Female adult (alcohol preserved, skull), MHNG 1971.067 (field collection number MZ261). Mozambique, Zambezia Province, Mount Namuli, camp on mountain plateau, 15.405933°S, 37.067006°E, ca. 1,200 ma.s.l., 20 November 2008. GoogleMaps Mist netted over river in fragment of mid- altitude forest on mountain plateau, ca. 18:45 h. Reference sequences of the mitochondrial CYT-B and COI genes issued from these paratypes are deposited in the GenBank under accession numbers MZ936305 View Materials – MZ936308 View Materials and MZ936309 View Materials – MZ936312 View Materials , respectively GoogleMaps .
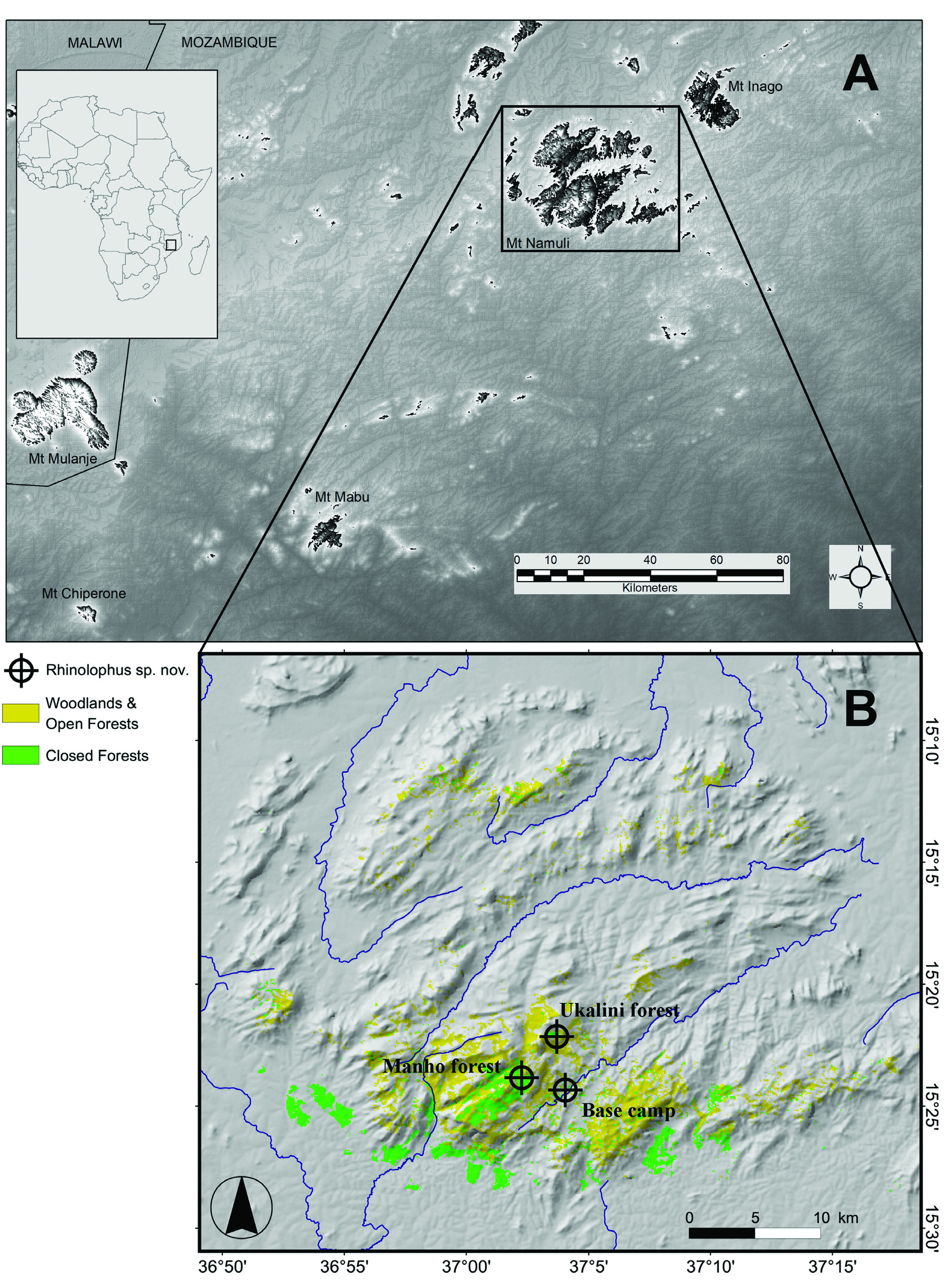
Referred material.— Female adult (alcohol preserved, skull), DM 10839 . Mozambique, Zambezia Province, Mount Namuli, Manho Plateau Forest , 15.3975°S, 37.0372°E, ca. 1,734 ma.s.l., 20 November 2008. Mist netted over small stream in forest clearing at approximately 18:30 h. GoogleMaps Male adult (alcohol preserved, skull, baculum preserved in glycerol), DM 10833 (field collection number MZ099). Mount Namuli, base camp on mountain plateau, 15.405933°S, 37.067006°E, ca. 1,200 m a.s.l., 20 November 2008. Mist netted over river in remnant fragment of mid- altitude forest on mountain plateau, ca. 18:00 h. GoogleMaps Other referred material are five specimens previously reported as R. cf. denti / simulator by Demos et al. (2019), which also proved to belong to R. namuli sp. nov. based on very similar CYT-B lineages. These specimens were collected approximatively 8 km north and 8 km east of Gurue , Mozambique ( FMNH 17708 – 177110 ), GoogleMaps in the Zovo Chipolo Forest, Nyika National Park, Malawi ( FMNH 191585 ), GoogleMaps and in the Ndundulu Forest, 9 km east of Udekwa Udzungwa Mts , in GoogleMaps Tanzania .
Type locality and distribution.— Mount Namuli (15.369250°S, 37.061361°E, ca. 1,650 ma.s.l.), ca. 10 km north-west of Gurue , Zambezia Province, north-central Mozambique (central Mozambique). Recorded in montane evergreen forest and nearby forest clearings on the high plateau and below the peak of the mountain at altitudes ranging 1,220 to 1,650 m GoogleMaps .
Etymology
The species epithet namuli is a noun in apposition and is a derivative of the type locality Mount Namuli, Zambezia Province, Mozambique.
Nomenclatural statement.—A life number was obtained for the new species Rhinolophus namuli : urn:lsid:zoobank.org:act:7A9806D9-CCC6-41A2- B898-6DCDEFFFBB51.
Diagnosis
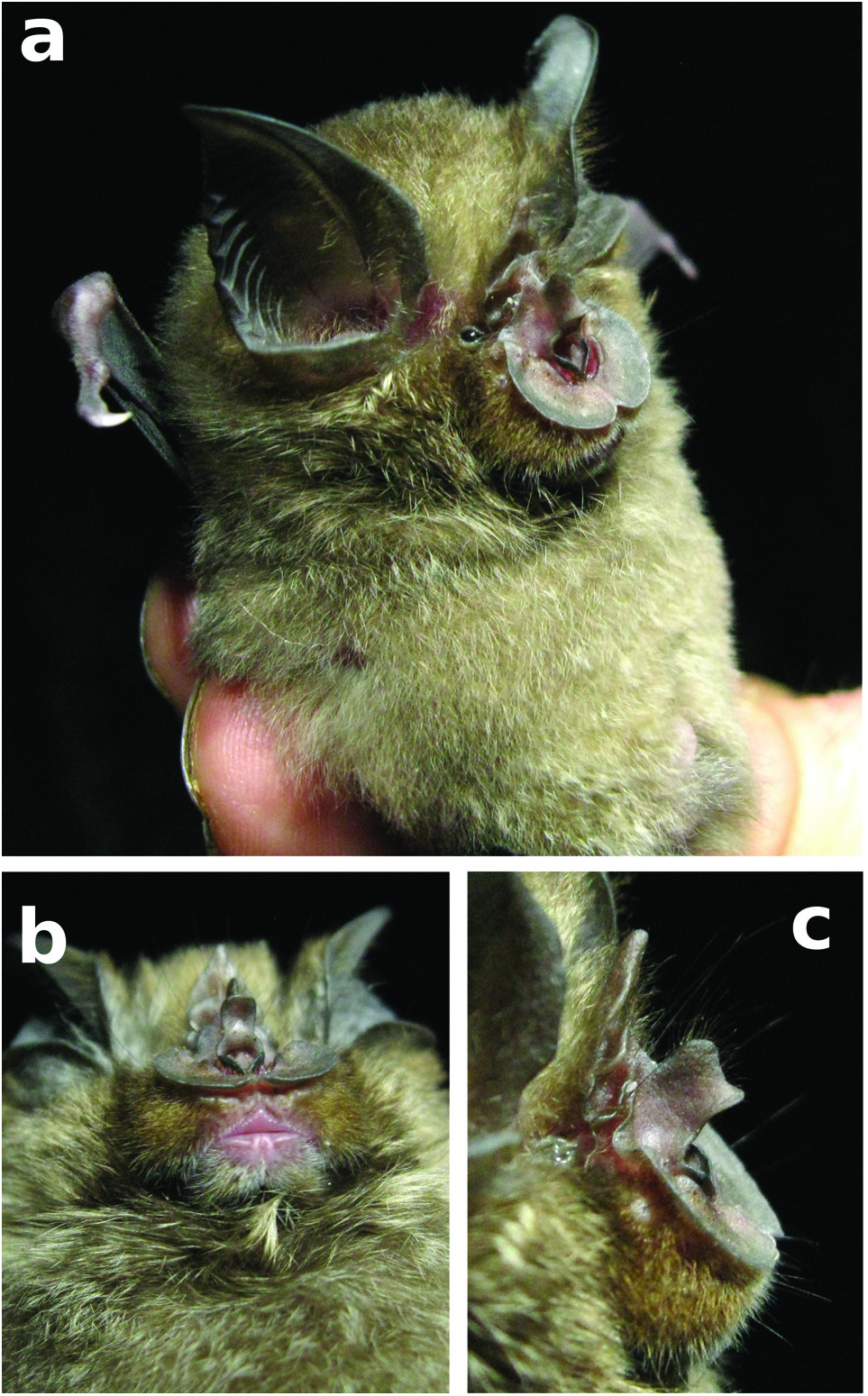
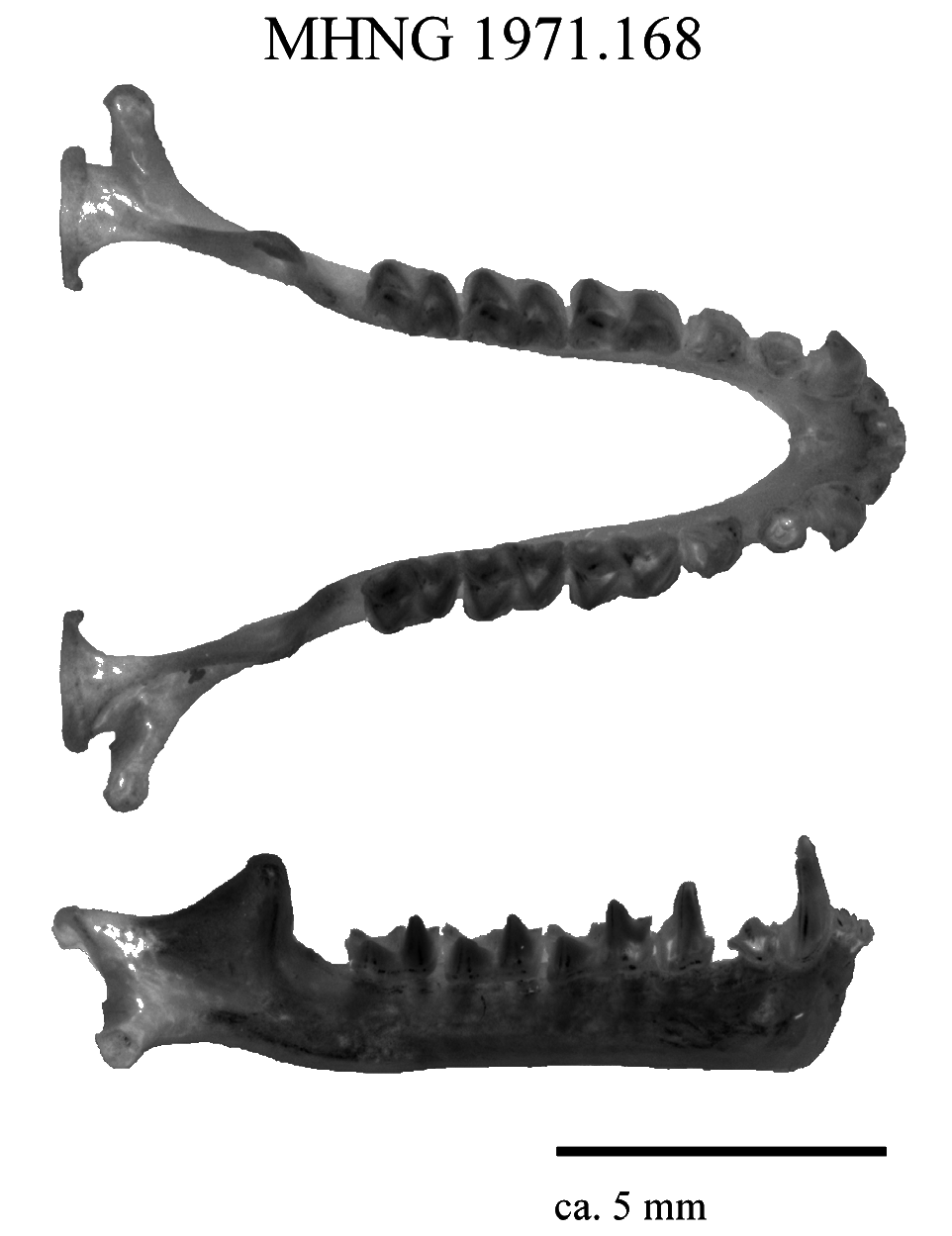
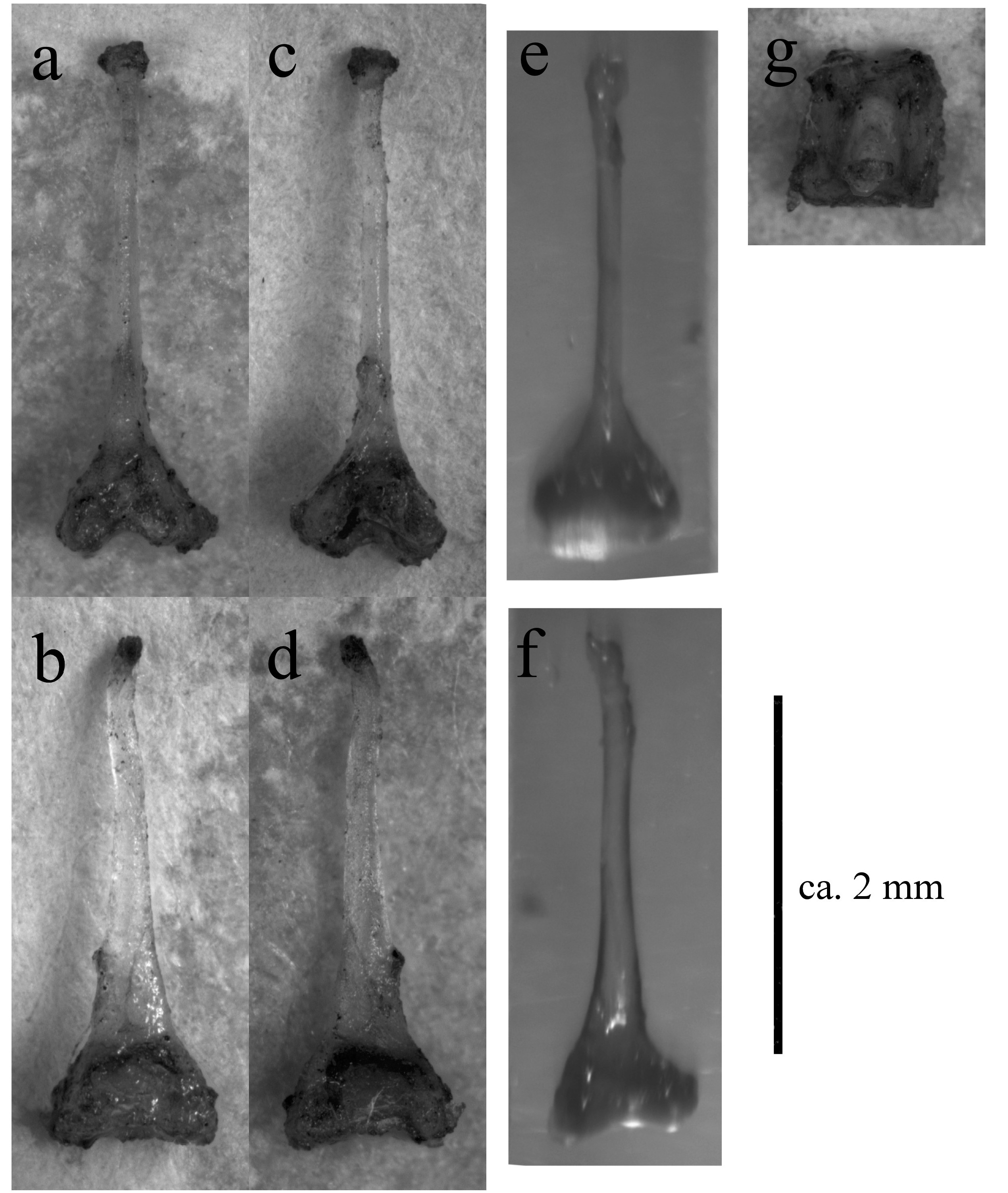
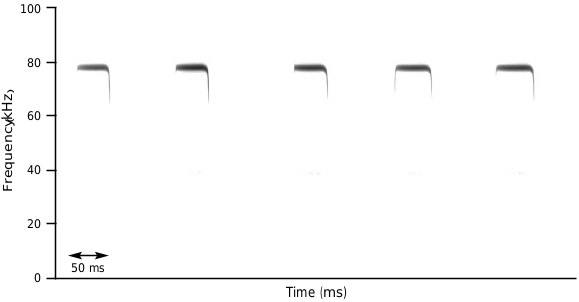
Medium sized bat (forearm length 47.2–49.5 mm) with medium sized ears and three well-defined mental groves on lower lip (Supplementary Appendix S2 Fig. A1 View FIG ). Horseshoe and sella wide ( Fig. 4 View FIG ), lancet concave with a narrow tip (hastate), connecting process high and rounded, with scattered hairs present along its ridge ( Fig. 5 View FIG , Supplementary Appendix S2 Fig. A2 and View FIG A3). Elongated first phalanx of third digit relative to metacarpal (Table 3). Bony bar forming the infraorbital foramen is present and slender, inflated nasal swellings ( Fig. 6 and View FIG Supplementary Appendix S2 Fig. A4 View FIG ), length of palatal bridge 36% of upper toothrow length (Table 4). Minute pm3 either present or absent in the lower toothrow, with pm2 and pm4 not making contact ( Fig. 7 View FIG ). Baculum medium-sized, dorsal and ventral parts of the basal cone subequal in size with dorsal and ventral incision shallow ( Fig. 8 View FIG ). In lateral view, baculum shaft narrows towards tip, undulating in shape and forms an evenly rounded ventrally bent tip. In dorsoventral view, shaft straight with shallow constriction at inflection point of spatulated tip (Supplementary Appendix S2 Fig. A5 View FIG ). Rhinolophus namuli sp. nov. echolocates at a mean peak frequency of 76.9 kHz, SD = 0.8, range = 75.9–78.0 ( Fig. 9 and View FIG Table 5). Phylogenetically, this species belongs to the capensis -group.
Description
Dorsal pelage near uniform beige brown, slightly darker towards head, turning near rusty on both sides of the muzzle. Ventral fur paler than dorsal. Hair length ca. 7 mm, with dark base becoming lighter towards tips causing slight shimmering effect. At time of capture, both male and female live animals exhibited musky and somewhat unpleasant smell.
Head.—Ears medium sized, just reaching tip of muzzle when laid forward, tips pointed in holotype but bluntly rounded in both paratypes, 8–9 ear folds. Three well-developed mental grooves on the lower lip, both lips devoid of hair ( Fig. 4 and View FIG Supplementary Appendix S2 Fig. A1 View FIG ). The mental grooves were diagnosed according to Csorba et al. (2003: Fig. vi, p. xxviii) and are shown in close-up in Supplementary Appendix S2 ( Fig. A1 View FIG ). Noseleaf completely covers muzzle, horseshoe wide and secondary noseleaf absent. Sella naked, wide at base, slightly constricted in the middle but otherwise almost parallel sided with rounded tip ( Fig. 5 and View FIG Supplementary Appendix S2 Fig. A2 View FIG ). Internarial cup rather well developed. Internarial septum is not markedly expanded. Connecting process somewhat hairy, high and rounded, basal part rises near parallel to sella. Tip of the lancet is hastate, with concave sides and a pointed, tapering tip. The hastate lancet differentiates R. namuli sp. nov. from R. maendeleo and other members of the adami -group ( Fig. 5 and View FIG Supplementary Appendix S2 Fig. A2 View FIG ), more closely resembling members of the capensis -group (see Supplementary Appendix S2 Fig. A3 View FIG for a comparison of lancet shape with R. simulator ).
Wing bones.—Metacarpal of the 4th and 5th digit subequal in length, always longer than the 3rd digit (Table 3). Length of the 1st phalanx greatest in 3rd digit, followed by 5th, then 4th digit. Length of the 2nd phalanx greatest in 3rd digit, followed by 4th, then 5th digit. 2nd phalanx of 4th digit greater than twice the length of 1st phalanx.
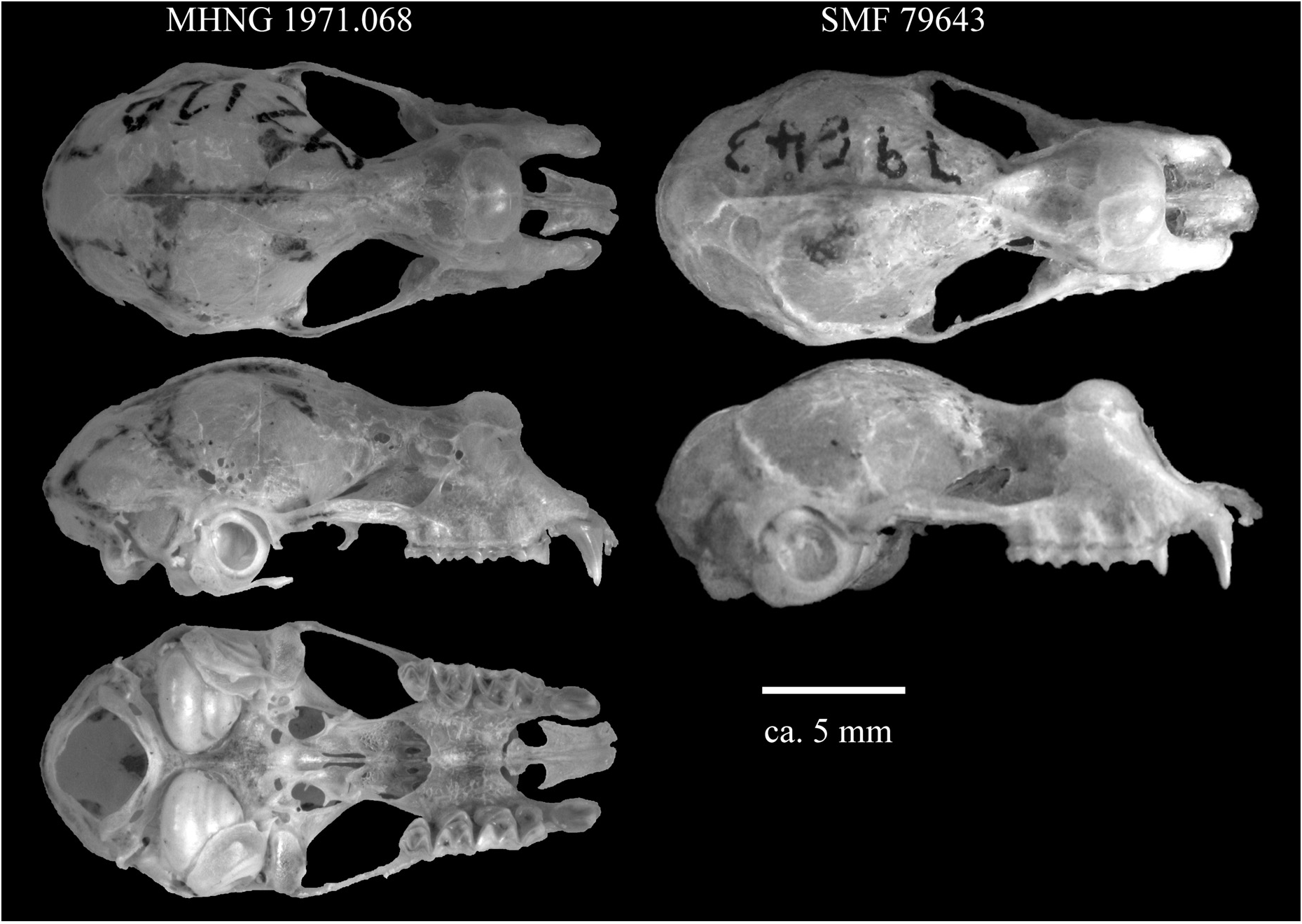
Cranium.—Skull is narrow (mastoid width exceeds zygomatic width) in MHNG 1971.068 (holotype, male), MHNG 1971.069 (paratype, female), and DM 108333 (referred material, male), but robust (zygomatic width exceeds mastoid width) in MHNG 1971.067 (♁) and DM 10839 (referred material, ♀). Differences are slight, indicating within-species variation in skull shape (Table 4). Weakly developed sagittal crest extends roughly three quarters along the length of the braincase from well-developed supraorbital crest (sagittal crest equally developed in both male and female). Posterior-most point of skull (meeting point of lambdoid and sagittal crest) positioned markedly above the lateral plane formed by the zygomatic arches. Anterior nasal swelling inflated, moderately wide and high, angled forward and protruding anteriorly in lateral view ( Fig. 6 and View FIG Supplementary Appendix S2 Fig. A4 View FIG ). Anterior median and lateral nasal swellings well-developed but posterior median swellings deflated. Moderate rostral depression between supraorbital ridges creates a concave rostral profile. Posterior-most point of rostral depression reaches, but does not extend beyond, constriction point between the orbits ( Fig. 6 View FIG ). The infraorbital foramena are enclosed by a bony bar on both sides in all specimens collected, which is illustrated in close-up in Supplementary Appendix S2 ( Fig. A4 View FIG ). Note that in specimen DM 10833, initial inspection concluded the bar was absent on the left side (as reported by Monadjem et al., 2010 a), but subsequent investigation confirms it presence on both sides (L. Richards, personal communication). Length of the palatal bridge moderate, about one third CM3 length (34.1–37.3%).
Teeth. — Dental formula I 1 /2, C 1/1, PM 2/2, M 3/3. Upper canines moderately well developed, extending about twice the length of PM4 ( Fig. 6 View FIG ). PM2 small, slightly extruded, separating C1 and PM4 and present on both sides. Minute pm3 present on left side and partly hidden by crown of pm2, lacking on right side, with pm2 and pm4 separate on both sides ( Fig. 7 View FIG ).
Baculum.—The baculum is medium-sized with a total baculum length of 2.85 mm (range ca. 2.8– 2.95 mm), greatest lateral width of basal cone of 0.78 mm and greatest dorsoventral width of basal cone of 0.87 mm ( Fig. 8 and View FIG Supplementary Appendix S2 Fig. A5 View FIG ). The basal cone occupies about 20% of the total length, with dorsal and ventral parts subequal in size, neither exhibiting a relative proximally projection. Both ventral and dorsal incisions of the basal cone are shallow. In lateral view, the baculum shaft constricts gradually, before reaching a constant width at about half the length to the tip. The shaft is not straight, but rather undulating. The tip is evenly rounded and distinctively curves ventrally at an angle of about 30° from the proximaldistal axis. In dorsoventral view, the shaft extends from the basal cone largely straight, unflattened and of equal width almost along its entire length. At the inflection point where the tip begins to curve ventrally (about 4/5th along the total baculum length), there is a shallow constriction in the shaft when viewed from a dorsoventral perspective. The tip thus appears spatulated and rounded ( Fig. 8 and View FIG Supplementary Appendix S2 Fig. A5 View FIG ).
Echolocation.—Calls recorded from seven individuals had a mean peak frequency of 76.9 kHz and a range of 75.9–78.0 kHz ( Fig. 9 and View FIG Table 5).
Comparisons
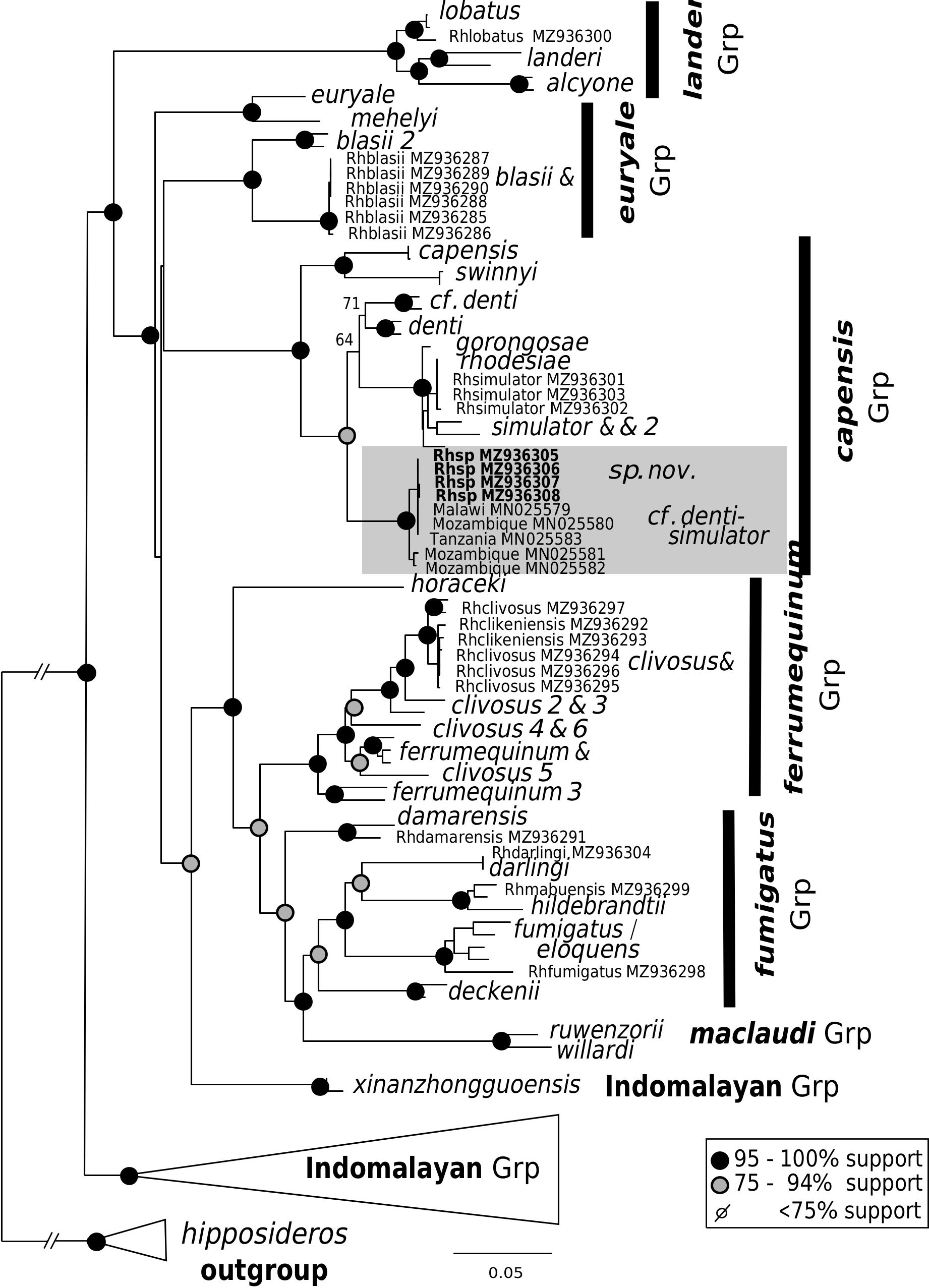
Taxonomic position.—The connecting process is well-developed, high and rounded, but is neither continuously arched (pearsoni -group, ferrumequinum -group or fumigatus -group), low and rounded ( hipposideros -group), low and poorly developed (maclaudi -group) nor sharply triangular ( landeri -group) ( Csorba et al., 2003). The sella is naked and constricted in the middle rather than hairy ( fumigatus -group) and/or parallel-sided (euryale - group). Three mental groves are well developed (Supplementary Appendix S2 Fig. A1 View FIG ), not partly obliterated like in members of capensis -group. These features, in addition to the characters described above, indicate R. namuli sp. nov. bears most resemblance morphologically to R. maendeleo , which was moved, with R. adami , from the capensis -group into a newly created adami -group by Kock et al. (2000). Regarding the defining characters of the adami -group, R. namuli sp. nov. has a large sella, straight-sided or rounded lancet, three well-defined mental grooves, small second premolar in the toothrow and more bulbous narial swellings. The nasal swellings are pronounced and clearly align to species of the adami - rather than capensis -group when R. namuli sp. nov., R. capensis , R. simulator and R. adami are compared side-by-side (see Supplementary Appendix S2 Fig. A6 View FIG ). However, the ear length of R. namuli sp. nov. is medium rather than large, only three of the five specimens of R. namuli sp. nov. have a narrow skull, and all have a moderate palate (36% of CM 3 in namuli sp. nov. against> 40% in adami -group). Furthermore, when viewed in profile, namuli sp. nov. has a high and rounded connecting process, which is lower and regularly arched in the adami -group ( Csorba et al., 2003). Morphologically, Rnamuli sp. nov. should therefore be retained within the capensis -group. Our genetic analysis confirms this, with a robust positioning of Rnamuli sp. nov. in the capensis -group alongside R. simulator , R. swinnyi and R. denti as closest relatives ( Fig. 3 View FIG ). Unfortunately, as no genetic sequences for R. maendeleo or R. adami are currently available, we cannot assess formally their phylogenetic position within the Rhinolophidae ra - diation. We therefore leave open the question of the validity of the adami -group and whether the striking morphological similarities shared between maendeleo and namuli sp. nov. represent convergences or true synapomorphies.
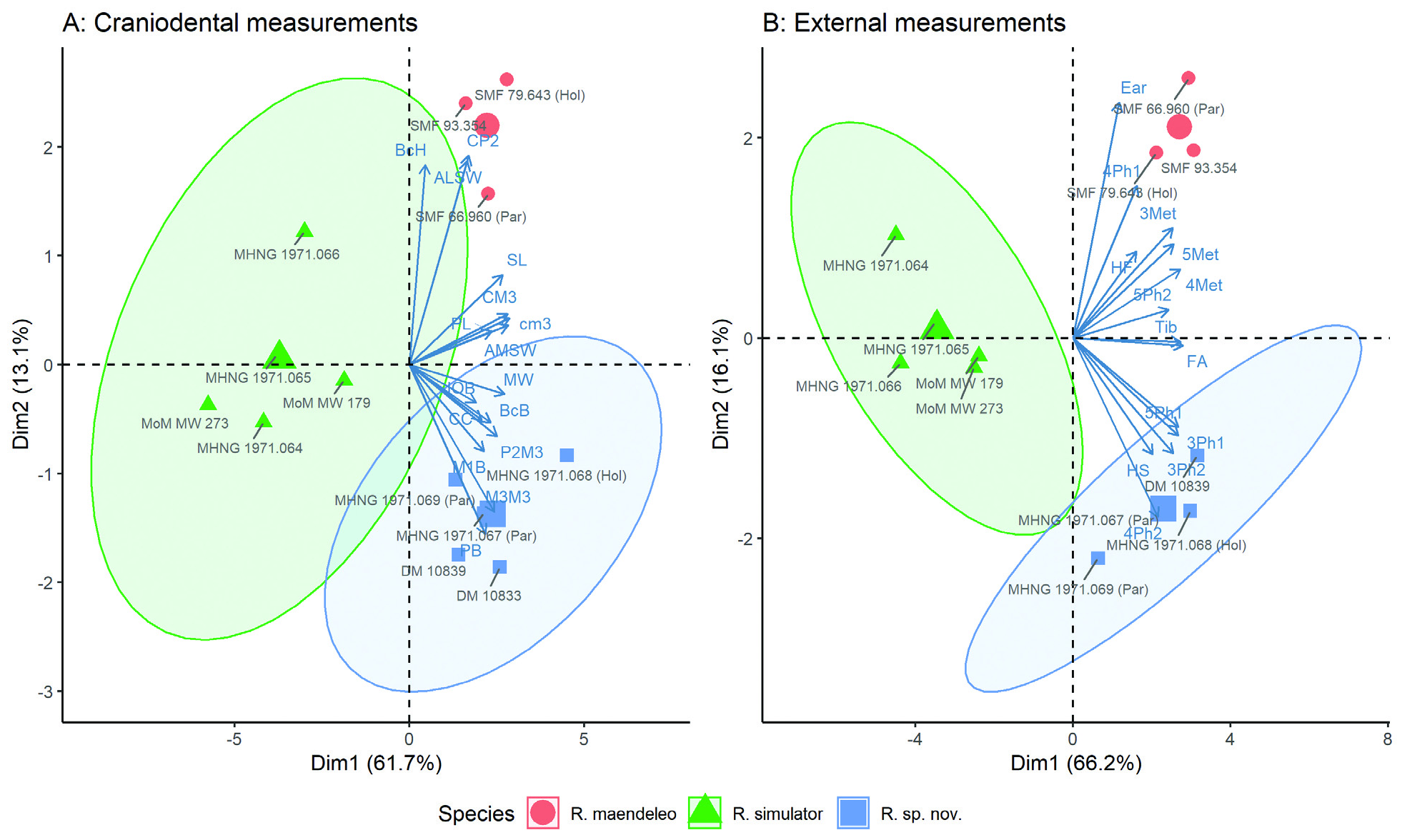
Specific status.— Rhinolophus namuli sp. nov. appears morphologically most similar to R. maendeleo , although these two species are currently placed in two separate groups ( capensis -group and adami -group, respectively). Due to this mismatch, R. maendeleo is used as the primary comparison to assign specific status, while other members of the adami and capensis groups are considered under ‘Similar species’ below. In comparison to R. maendeleo , R. namuli sp. nov. shares several similar external and craniodental measurements, including overlapping forearm length, horseshoe width, metacarpal length of 4th and 5th digit, length of 1st phalanx of 3rd and 5th digit, length of 2nd phalanx of 5th digit, Tib, HF, MW, ZW, BcH, CC, CM3, IOB, ML, cm3, AMSW, PL (Table 3). The two species also exhibit a similar overall cranial shape and size ( Fig. 6 View FIG ). However, R. namuli sp. nov. is distinguished from R. maendeleo by a number of key characters. The characteristic feature of R. maendeleo , an absence of the bony bars forming the infraorbital foramena ( Kock et al., 2000), is not shared in R. namuli sp. nov, where the bony bars forming the infraorbital foramena are present in all collected specimens (Supplementary Appendix S2 Fig. A4 View FIG ). R. namuli sp. nov. can also be distinguished from R. maendeleo by a number of non-overlapping craniodental measurements that indicate a slightly shorter overall skull length (SL — Table 4), a slightly wider and shorter rostrum (M3M3 and PB — Table 4) and subtle differences in rostral and narial inflation shape ( Fig. 6 View FIG ). R. maendeleo also bears a deeper and longer rostral depression whose posterior most point extends beyond the constriction point between the orbits. In R. namuli sp. nov. the depression is shallower, and does not extend as far back between the orbits ( Fig. 6 View FIG ). These differences in rostrum shape, inflation angle and depression are captured in the two angle measurements (Ang1, Ang2), of which the first is non-overlapping between the species (Table 4).
The two species also differ completely in bacular morphology ( Fig. 8 and View FIG Supplementary Appendix S2 Fig. A5 View FIG ). In lateral view, the baculum of R. maendeleo narrows gradually from the basal cone to the tip, the shaft being straight and flattened dorsoventrally, especially towards the distal end ( Kock et al., 2000). In contrast, the baculum of R. namuli sp. nov. is undulating, uncompressed, and is distinctively downturned (ventrally angled) at the tip. The infla - tions forming the basal cone of the baculum of R. maendeleo are unequal with the dorsal inflations protruding proximally relative to the ventral. This is in contrast to R. namuli sp. nov., which bears subequal inflations with no clear protrusion. The species also differ in the extent of the ventral incision in the basal cone, which is deep in R. maendeleo and shallow in R. namuli sp. nov. (Supplementary Appendix S2 Fig. A5 View FIG ).
Externally, the two species are distinguished by the length of the tail, size of the ears and differences in horseshoe structure, and in the dimensions of the wing bones (Table 3). At a mean length of 25.9 mm, the tail of R. maendeleo is markedly longer than that of R. namuli sp. nov. (mean of 20.3 mm). Ear length follows a similar pattern (means of 23.5 mm versus 19.3 mm in R. maendeleo and R. namuli sp. nov., respectively). The connecting process of R. namuli sp. nov. is high and rounded, not continuously arched as in R. maendeleo , and with a more pronounced constriction in the sella ( Figs. 4 and View FIG 5, and Supplementary Appendix S2 Fig. A2 View FIG ). Within the wing bones, measurements of metacarpal and phalanx length illustrate differences between the species, particularly in the 3rd and 4th digits (Table 3).
Similar species. — Kock et al. (2000) compare R. maendeleo to the morphologically similar R. adami , illustrating a set of characters that differ between the species. We confirmed that R. namuli sp. nov. possesses many of the same distinguishing characters. Specifically, both R. maendeleo and R. namuli sp. nov. can be distinguished from R. adami by a smaller and concave-sided lancet, a less arched connecting process, a near-parallel sided interpterygoid groove, differing bacular morphologies, external and craniodental measurements, including palate length and inflation breadth ( Kock et al., 2000).
When compared to members of the capensis - group, R. namuli sp. nov. is larger than R. simulator , with non-overlapping forearm, tibia and wing bone lengths (Table 3) and several non-overlapping craniodental measurements, such as greatest skull length, mastoid width, zygomatic width and lower toothrow length (Table 4). Based on a comparison of published measurements of R. denti , R. swinnyi , R. gorongosae and R. rhodesiae, Rnamuli sp. nov. is much larger and more robust as highlighted by non-overlapping forearm length and greatest skull length. It also echolocates at a much lower frequency that any of these species (Table 2). R. namuli sp. nov. has more inflated anterior nasal swellings than members of the capensis -group (Supplementary Appendix S2 Fig. A6 View FIG ). Finally, the bacular morphology of R. namuli sp. nov. sets the species apart from all potentially similar species from the adami and capensis groups to which it bears closest morphological affinity.
Summarizing the differences highlighted above, the PCA results comparing R. namuli sp. nov. to both R. maendeleo and R. simulator (the two most similar species from the adami and capensis groups, respectively) illustrate the distinct position of the three species ( Fig. 2 View FIG ). While 95% confidence intervals (CIs) for the first two PCs overlap slightly between R. namuli sp. nov. and R. simulator , such uncertainty is to be expected given the small sample size. For R. maendeleo , the sample size was too small to estimate CIs. The first two PCs explained a large amount of variance in the data, with a cumulative 76% for craniodental measurements and 82.2% for external measurements.
Phylogenetic reconstructions based on the CYT-B gene ( Fig. 3 View FIG ) demonstrate that R. namuli sp. nov. belong to the capensis -group with strong support. Compared to the closely related and sympatric R. simulator , R. namuli sp. nov. is genetically distinct, confirming the morphological differences presented above. In the future, additional studies on the phylogenetic relationships between the species of the adami - and capensis -group would be desirable to ascertain relationships or test their taxonomic validity.
Rhinolophus namuli sp. nov. echolocates at a frequency that is not shared by other species in the region (Table 5). At a mean frequency of 76.9 kHz, the calls of R. namuli sp. nov. differs markedly from R. simulator (86 kHz), R. clivosus (80.5–81.3 kHz) and R. fumigatus (58.4 kHz), the three most similarly echolocating species in our dataset for the region (Table 5).

| MHNG |
Switzerland, Geneva, Museum d'Histoire Naturelle |
| FMNH |
USA, Illinois, Chicago, Field Museum of Natural History (also used by Finnish Museum of Natural History) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Rhinolophus namuli sp. nov. Curran, Kopp, Ruedi, and Bayliss, 2022
| Curran, Michael, Kopp, Mirjam, Ruedi, Manuel & Bayliss, Julian 2022 |
Rhinolophus cf. denti
| Thomas 1904 |
Rhinolophidae
| Gray 1825 |