
Placosoma limaverdorum, Borges-Nojosa, Diva Maria, Caramaschi, Ulisses & Rodrigues, Miguel Trefaut, 2016
|
publication ID |
https://doi.org/10.11646/zootaxa.4169.1.8 |
|
publication LSID |
lsid:zoobank.org:pub:B54A654B-D5FA-4707-9C31-BAFAF98D5F53 |
|
DOI |
https://doi.org/10.5281/zenodo.5680789 |
|
persistent identifier |
https://treatment.plazi.org/id/03AB9335-2A24-FFF4-C3FC-F8C24A20599E |
|
treatment provided by |
Plazi |
|
scientific name |
Placosoma limaverdorum |
| status |
sp. nov. |
Placosoma limaverdorum sp. nov.
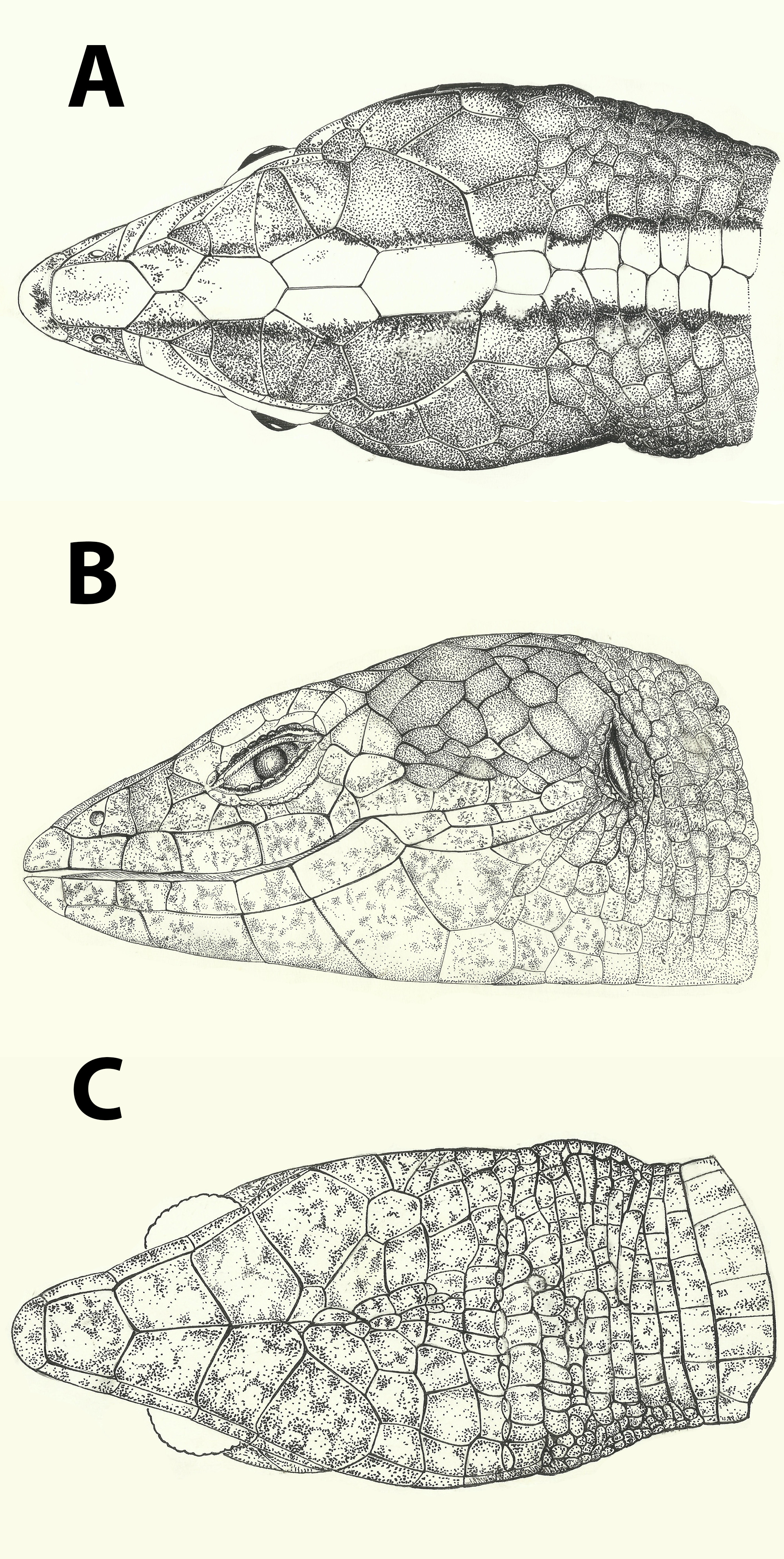
( Figures 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
Holotype. CHUFC L 1998, an adult male, collected at the Maciço de Baturité (Sítio Olho d’Água dos Tangarás), Pacoti Municipality, state of Ceará, Brazil (04o14’9,41”S / 38o55'1,15"W; Datum WGS 84) by J.A. Carneiro in December, 1996 ( Figures 1–2 View FIGURE 1 View FIGURE 2 ). GoogleMaps
Paratypes. CHUFC L1958, juvenile male, November 15, 1995, and MZUSP 88071 (CHUFC L2055), adult male, February 19, 1998, collected by D.M. Borges-Nojosa; CHUFC L2527, adult male, CHUFC L2525 and CHUFC L2526, adult females, August 27, 1998, collected by D.C. Lima; CHUFC L3116, adult female, December 12, 2003, collected by Y. Quinet; CHUFC L4170 ( Figure 3 View FIGURE 3 ), adult male, and CHUFC L4171 ( Figure 3 View FIGURE 3 ), adult female, June 26, 2010, collected by D.C. Lima and T.A. Sousa; CHUFC L4194, adult male, August 22, 2010, collected by D.C. Lima; MNRJ 26049 (CHUFC L5649), adult male, September 27, 2014, MPEG 32210 View Materials ( CHUFC L5783), juvenile male, October 18, 2014, collected by D.M. Borges-Nojosa, D.P. Castro et al.: all from Serra de Maranguape , Maranguape Municipality, State of Ceará, Brazil (03o54'– 04o03' S / 38o32'– 38o40' W; Datum WGS 84) GoogleMaps . CHUFC L3720, juvenile male, March–July 2005, collected by L.B.M. Brito: Maciço de Baturité , Guaramiranga Municipality (Sítio Guaramiranga), State of Ceará, Brazil (04o05– 04o40’ S / 38o30'– 39o10' W; Datum WGS 84). GoogleMaps
Etymology. The species honors Prof. Dr. José Santiago Lima-Verde, an important collaborator of herpetological studies in the State of Ceará, who started the Herpetological Collection of the Universidade Federal do Ceará (CHUFC) and the Núcleo Regional de Ofiologia da UFC (NUROF-UFC). This description is also a tribute to the agronomist Wilson Luiz Lima-Verde, owner of the cottage (Sítio Olho d’Água dos Tangarás) where the holotype was obtained.
Diagnosis. Body long, depressed; head long with a pronounced acute snout; tail longer than body; tongue elongate, arrow headed. Limbs slender, pentadactyl, all clawed. Ear opening and eyelid distinct, with an undivided transparent palpebral disk. Frontonasal single; prefrontals, frontoparietals, parietals, interparietal and two series of occipitals present; four supraoculars. Nostril in the center of a divided nasal. Tympanic recess deep. Collar absent, gular fold present; four enlarged pairs of chinshields, posteriormost reduced. Dorsal scales smooth, subimbricate, juxtaposed, disposed in regular transverse rows. Lateral scales distinct in shape and much smaller than dorsals and ventrals, disposed in irregular transverse rows. Ventrals quadrangular, smooth, imbricate, larger than dorsals, disposed in regular transverse and longitudinal rows. Males with femoral pores, preanal pores absent; females without femoral pores.
The systematics of gymnophthalmid lizards is going through a period of effervescence since the advent of the first comprehensive molecular study showed that morphologic character convergence was widespread, introducing uncertainty in their taxonomy ( Pellegrino et al. 2001). Since then, several genera and suprageneric groups have been defined molecularly but we have no consistent morphological synapomorphies supporting either the present admitted genera and tribes ( Rodrigues et al. 2009, Torres-Carvajal et al. 2016, Goicoechea et al. 2016). The genus Placosoma has been consistently recovered in molecular studies, nested among the Cercosaurini with high support ( Pellegrino et al. 2001, Kok 2015, Torres-Carvajal et al. 2016, Goicoechea et al. 2016). Nevertheless, no morphological synapomorphies have been proposed either to the genus or to the tribe Cercosaurini. Considering the absence of molecular data for the species herein described and while waiting for a sound morphological study of the entire tribe to better characterize its genera we use the following character combination to support the inclusion of the new species in the Cercosaurini and in the genus Placosoma : presence of an eyelid and ear opening, a long body and tail, pentadactyl limbs, presence of an elongate and acute snout, two series of occipital scales, homogeneous and slightly imbricate quadrangular dorsal and ventral scales, lateral scales distinct in size and shape from dorsal and ventral ones but never granular. The genus Placosoma differs from the stream adapted genera Echinosaura , Neusticurus , Potamites , and Gelanosaurus by having a homogeneous dorsal scalation (heterogeneous in size and shape and ornamentation in those genera). From Cercosaura and Pantodactylus it differs by presenting eight longitudinal series of quadrangular scales at midbody and no granular scales in the flanks (six or less enlarged rows of ventrals and granular scales on flanks). From Anadia , Euspondylus , Macropholidus , and Riama it can be distinguished by having a series of lateral scales smaller than dorsal and ventral scales but never granular (generally identical to dorsals, without distinction). From Proctoporus , Petracola and Pholidobolus it differs by the absence of a lateral fold with granular scales on sides of body (present).
Placosoma limaverdorum differs from all congeners by having an undivided transparent palpebral disc (divided with scales in all other species), an entirely divided nasal with a nostril in the center, a deeply tympanic recess, absence of preanal pores, and by the presence of a light vertebral stripe extending from frontonasal to tip of tail, with about one scale wide, bordered laterally by a dark pigmentation, irregularly interrupted by interspaced ocelli (two paravertebral light stripes in P. cordylinum , P. champsonotus , and P. g l ab e l l um, and two paravertebral stripes and a lateral one in P. cipoense ). It further differs from P. glabellum by having squared ventral scales (posterior part of scales rounded), and from P. champsonotus by having smooth dorsal scales (keeled) and a maximum of six enlarged dorsal scales between fore limbs (eight or more). Placosoma limaverdorum share with P. glabellum , P. cordylinum and P. cipoense a low number of enlarged dorsal scales between forelimbs (maximum of six) but differ from those species in number of femoral pores 21–26 (19–22 in P. glabellum , 27–32 in P. cordylinum and 26 in P. cipoense ). Placosoma cipoense is the only species in the genus to have the superior part of nasal semidivided; the suture in also present in the lower part of nasal in P. limaverdorum . Additionally, in P. limaverdorum , the frontal scale has anterior and posteriorly the same approximate width, is smaller than parietal and internasal and is either totally separated from the first supraocular or in punctual contact with it. In all other species the frontal scale is longer than parietal and internasal and is in broad contact with the first supraocular. Another difference is that the suture between frontoparietals is relatively longer in P. limaverdorum when compared with those of its congeners.
Description of the Holotype. Measurements (mm): snout-vent length 52,8; tail length 60,2 (regenerated); head length 13,4, head width 8,2 and head height 7,0; right forelimb and hindlimb,13,8 and 20,5, respectively. Snout long and pointed, neck distinct, with a conspicuous gular fold, collar fold absent. Rostral broad, visible dorsally, wider than high, in contact with frontonasal, nasal and first supralabial. Frontonasal pentagonal, long, in lateral contact with the nasal, indenting posteriorly a pair of prefrontals. Frontal hexagonal, longer than wide, as long as frontonasal and frontoparietal suture, contacting laterally first (punctualy), second (largely) and third (moderately) supraoculars and posteriorly a pair of longer than wide pentagonal frontoparietals. Interparietal as wide as and longer than frontal, posteriorly rounded, indenting anteriorly the frontoparietals. Parietals wide, irregularly hexagonal, rounded laterally, contacting anteriorly fourth supraocular and frontoparietals, laterally temporals and posteriorly the occipitals. A row of three occipitals following parietal and interparietal the central one smaller the lateral ones in contact with the temporal and a posterior pair of nucals. Supraoculars four, first the smallest, the other three irregular in shape but with the same approximate size. Nasal large, divided above and below the median nostril, contacting inferiorly first and second supralabials. Loreal single, higher than wide, contacting posterior part of nasal, prefrontal, first superciliary, frenorbital, preocular and second supralabial. Frenorbital triangular, smaller than posterior part of nasal, in broad contact with first superciliar and preocular. Superciliaries six/five (R/L), first largest. Lower eyelid smooth, without scale division, with a transparent palpebral disc. Suboculars six/five (R/L), posterior three the largest. Postoculars one/two (R/L). Supralabials eight, 4th under the eye, 4th and 6th the largest. A series of temporals irregular in size and shape, those close to dorsal ones much enlarged. Ear opening semicircular bordered by series of small and flat elongated scales. Tympanum distinct, and close to head surface; external auditory meatus reduced. Mental small, semicircular, in straight contact with a pentagonal posmental which is as long as wide, wider posteriorly and in contact with first infralabial. Three enlarged pairs of genials, the first two in broad contact, the third pair separated by irregular scales in the middle, all contacting infralabials. A smaller pair of chinshields broadely separated at midline and by the infralabials by an elongated scale. Infralabials elongated, eight/seven (R/L), the latest very small, the fifth the longest. Dorsal region of the neck with a transverse row of two enlarged and wider than long nuchals at midline following occipitals. A series of six-seven pairs of midline dorsals, smooth, slightly smaller than the nucals following them and gradually becoming identical to other dorsals. Other dorsal scales quadrangular, with straight edges or slightly rounded, imbricate, smooth, and organized in 38 regular transverse rows (between nucals and posterior level of hindlimbs). Six enlarged dorsals between forelimbs. Lateral region with smaller scales, smooth, imbricate, elongated, not well defined in regular transverse rows. Gular region with 12 transverse rows of irregular scales between the last pair of chinshields and the interbrachial region (included). Interbrachial region with a distinct gular fold, with nine scales, all quadrangular, medial ones larger. A distinctive granular area near forelimb and hindlimb insertion. Ventral scales squared, smooth, slightly imbricate, organized in longitudinal rows and 23 regular transverse rows from the gular fold to the row before the upper preanals. Scales around midbody 33: 10 ventrals, 4+7 laterals, 12 dorsals. Preanal region with two series of smooth and almost juxtaposed scales. Upper row with four scales, central ones largest and wider than long, six scales in the lower row, all longer than large. Twenty one total femoral pores (10R/ 11L) separated by the preanal scales; preanal pores absent. Fore limbs with large, smooth, juxtaposed to slightly imbricate scales, except on ventral part of upper arm, where scales are much smaller, rounded, rhomboid, strongly imbricate. Anterior and ventral parts of hindlimbs with large, smooth, imbricate scales, identical to those on corresponding parts of forelimbs. Scales on posterior part of hindlimbs much smaller, becoming larger, imbricate on dorsal part of tibia. Carpal and tarsal scales large, imbricate, smooth. Supradigital lamellae smooth, imbricate. Palmar and plantar surfaces with smooth, small, tuberculate granules. Fingers and toes relatively long, clawed. Subdigital lamellae double proximally, single distally, 13/14 (R/L) on finger IV and 19/18 (R/L) on toe IV. Fingers and toes with the following relative sizes: 1 <2 = 5 <3 <4 and 1 <2 <3 <5 <4, respectively. Scales of tail smooth, longer than wide, rectangular, disposed in 45 transverse circular rows and in irregularly longitudinal series.
Coloring. Dorsal surfaces of body and tail and lateral part of tail grayish brown with a conspicuous light vertebral stripe, one and half to two scales wide, extending from rostral to the tail. Near the base of tail the light stripe widens assumes a greyish tonality and occupy most of its dorsal part. Vertebral stripe bordered on the sides along its entire length by dark colour, usually interspersed with irregularly disposed light ocelli, which also occur on the flanks and dorsal surfaces of the limbs. Ventral surface of head and body creamy white, dotted with small dark spots. Ventral parts of limbs and tail with the same general color but with less intense dotting. Lateral surface of the head with a short and thin light stripe extending from preocular to temporals and bordered above by a thinner dark stripe that cross ear opening, goes over the front limbs and extends to the flanks. Supralabials and infralabials light colored with scanty dark pigmentation.
Variation. There is no sexual variation in meristic or morphometric characters in the type series ( Table 1 View TABLE 1 ). Likewise, sexual color dimorphism is apparently absent although females seem more dimly colored than males. One male (CHUFC L4170) showed small orange spots disposed in a irregularly longitudinal series along sides of body, complemented by similar scattered spots on limbs, ventral region and near the cloaca ( Figure 3 View FIGURE 3 ). Preserved specimens become darker but maintain a colour pattern similar to that of the live specimens except for the intensity.
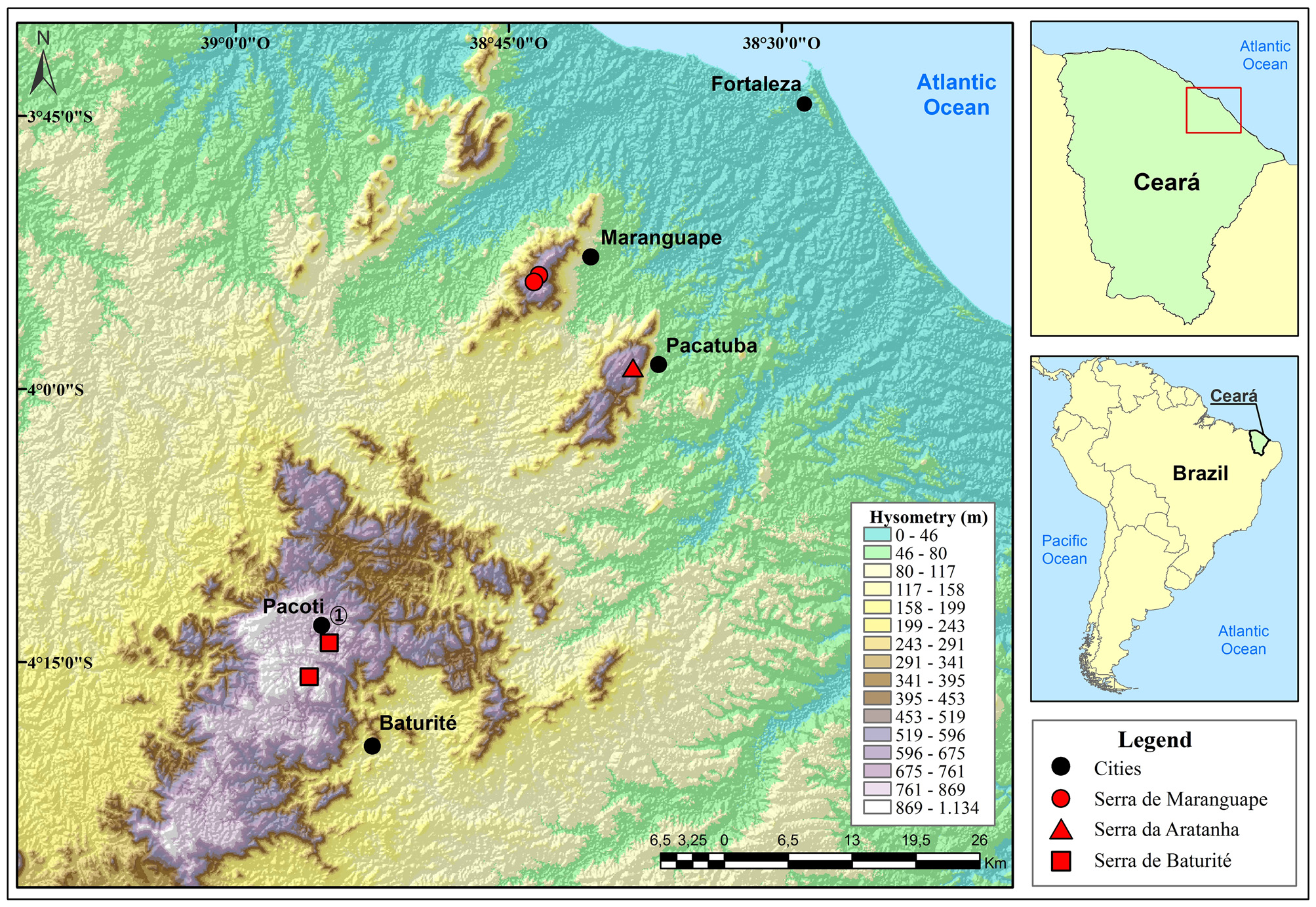
Distribution and natural history. The type series of Placosoma limaverdorum indicate that the species occurs in the forested mountains of Serra de Baturité (4o05’– 4o40’S /38o30’– 39o10’W), and Serra de Maranguape (3o54’– 4o03’S /38o32’– 38o40’W) in the state of Ceará, northeastern Brazil from where it is apparently endemic ( Figure 4 View FIGURE 4 ). Another record, although without voucher (P.C.M.D. Mesquita, personal information and photo) confirm its occurrence at Serra da Aratanha (3º55’– 3º58’S /38º38’– 38º36’W), another nearby forest region. These areas, the called “brejos de altitude”, are forest islands isolated in the semiarid Caatingas. Placosoma limaverdorum was always found inside the forest. Most specimens were spotted on the leaf litter of the primary and secondary vegetation, or in banana plantations ( Lima 2005), in sympatry with the gymnophthalmids Leposoma baturitensis Rodrigues & Borges, 1997 , Colobosauroides cearensis Cunha, Lima-Verde & Lima, 1991 and Stenolepis ridleyi Boulenger, 1887 . Two specimens were found in tree trunks, one on a central part of the trunk of a thin tree, about 1.3 m high, and the other in a vertical branch, about 1.2 m from the ground, between epiphytes and lichens. Three specimens were taken from the stomach contents of the snake Drymoluber dichrous ( Peters, 1863) from Serra de Maranguape ( Borges-Nojosa & Lima 2001).
Characters Holotype Paratypes Total (n=13)
Males (n=8) Females (n=4)


| MPEG |
Museu Paraense Emilio Goeldi |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.