
Neodiplopeltula, Holovachov & Boström, 2018
|
publication ID |
https://doi.org/10.5852/ejt.2018.458 |
|
publication LSID |
lsid:zoobank.org:pub:16C565AB-22A6-496F-AE95-0A876066F958 |
|
DOI |
https://doi.org/10.5281/zenodo.3843751 |
|
persistent identifier |
https://treatment.plazi.org/id/B1D9F161-1903-4227-8F97-5A2D6715F2CA |
|
taxon LSID |
lsid:zoobank.org:act:B1D9F161-1903-4227-8F97-5A2D6715F2CA |
|
treatment provided by |
Valdenar |
|
scientific name |
Neodiplopeltula |
| status |
gen. nov. |
Genus Neodiplopeltula gen. nov.
urn:lsid:zoobank.org:act:
Type species
Neodiplopeltula incisa ( Southern, 1914) gen. et comb. nov.
= Diplopeltis incisus Southern, 1914
= Diplopeltula incisa ( Southern, 1914)
= Diplopeltis incisus sensu Gerlach, 1950 partim (note #1)
= Diplopeltula incisa sensu Voronov, 1982 partim (note #2)
Diagnosis
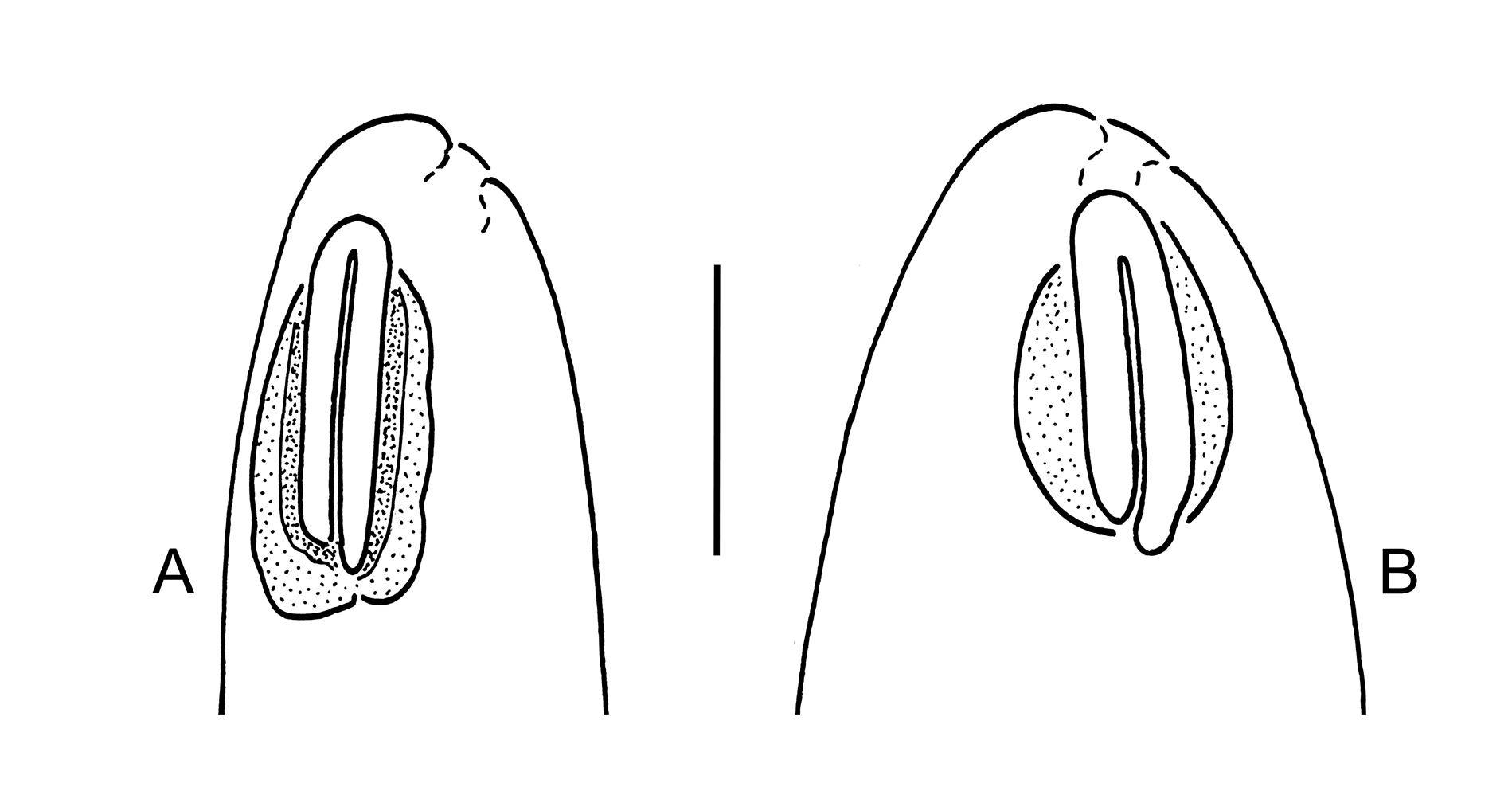
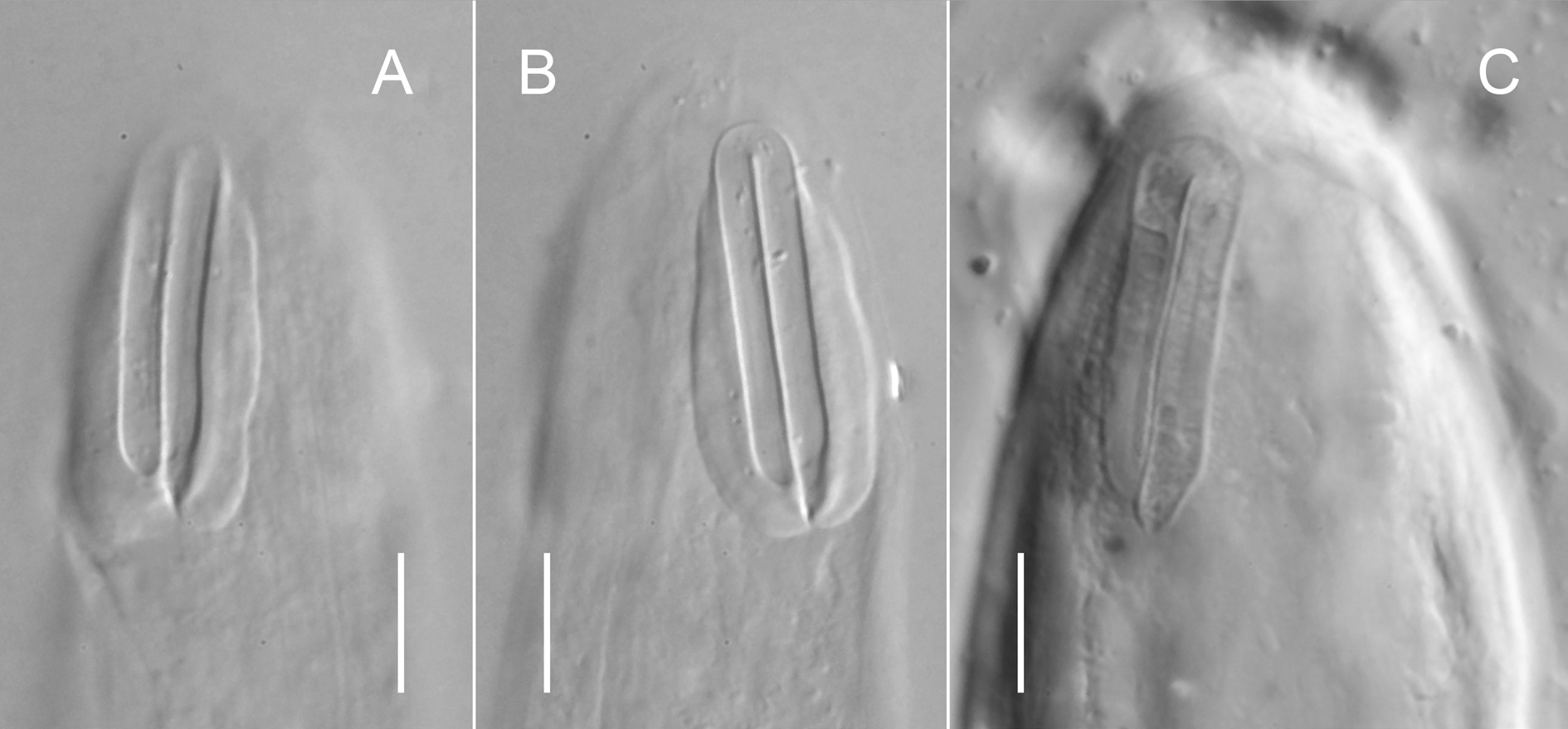
Cuticle transversely striated; striae visibly smooth under light microscope, may have fine longitudinal incisures visible under SEM. Lateral alae absent. Body pores and epidermal glands absent. Somatic sensilla present, most prominent along anterior part of pharyngeal region and on tail. Labial region bluntly rounded; lips fused. Six inner labial sensilla small, pore-like, located on anterior surface of lips, discernible under SEM only. Six outer labial sensilla papilliform, located on anterior surface of lips. Four cephalic sensilla setiform; their bases located at base of labial region, at level with anteriormost part of amphid. Bases of dorsosublateral setae often located more posteriorly than bases of ventrosublateral setae. Subcephalic sensilla absent. Cervical sensilla present in some species, setiform or papilliform, arranged in dorsosublateral and ventrosublateral rows at level with amphid and subcuticular periamphideal plates. Deirid and ocelli absent. Amphids in some species on strongly refractive lateral subcuticular periamphideal plates that can be connected together on ventral and dorsal sides forming a ‘cephalic framework’. Amphidial fovea loop-shaped (inverted U-shaped), dorsal branch often longer than ventral branch; both branches of amphideal fovea closely adjacent. Secretoryexcretory system present; renette cell located opposite to posterior part of pharynx or anterior part of intestine. Secretory-excretory ampulla present, located at level of pharynx. Cuticularised secretoryexcretory duct very short, opens to exterior either anterior or posterior to nerve ring level. Oral opening apical or shifted towards dorsal side of body. Buccal cavity usually well developed, subcylindrical and often asymmetrical: cheilostom usually broad, flexible; gymnostom subcylindrical, with weakly cuticularised walls; stegostom short conoid, its lining is uniform with lining of pharynx.Pharyngeal tubes absent. Pharynx subcylindrical, muscular, with evenly distributed myofilaments, gradually expanding towards posterior end; not subdivided into distinct sections; pharyngeal lumen uniform in thickness along entire pharynx length; valves absent. Cardia glandular, variable in shape and size, its posterior part embedded in intestinal tissue. Female reproductive system didelphic-amphidelphic, with equally developed branches, ovaries outstretched. Spermatheca present. Vulva equatorial or postquatorial. Vagina straight; pars proximalis vaginae encircled by large sphincter muscle; pars refringens vaginae absent, epiptygmata present or absent. Male reproductive system diorchic, testes opposed (anterior testis outstretched and posterior testis reflexed). Spicules symmetrical, arcuate; gubernaculum present, usually with strongly developed apophyses. Supplements, precloacal and postcloacal sensilla absent. Three caudal glands present, open via three separate openings, their cells and nuclei incaudal. Spinneret absent.
Etymology
The genus name is composed of a prefix ‘neo–’, meaning ‘new’ and Diplopeltula , and defines a new taxon proposed to accommodate species formerly placed in the genus Diplopeltula .
Relationships
The new genus differs from all other taxa currently included in the family Diplopeltidae (Holovachov 2017) in having amphids in the shape of an elongated loop, a well-developed subcylindrical stoma and outstretched ovaries. It is proposed in order to accommodate a number of species previously placed in the genus Diplopeltula and to solve a long lasting taxonomic conundrum ( Vincx & Gourbault 1992; Holovachov et al. 2009; Holovachov 2017; Holovachov & Boström 2017). The description of Diplopeltula breviceps , type species of the genus Diplopeltula , does not include clear descriptions of several important taxonomic characters (see Table 1 View Table 1 ) and the absence of such data prevents an unequivocal placement of the species D. breviceps and the genus Diplopeltula in the classification system of Nematoda. As a result, D. breviceps is considered species inquirenda et incertae sedis and the genus Diplopeltula is accordingly considered genus inquirendum et incertae sedis. The second species, D. longiceps , is also considered species inquirenda et incertae sedis for the same reasons (incomplete description and unclear systematic position).
Valid species
Neodiplopeltula barentsi ( Steiner, 1916) gen. et comb. nov.
= Diplopeltis barentsi Steiner, 1916
= Diplopeltis ovalis Ditlevsen, 1928 syn. nov. (note #3)
= Neodiplopeltula ovalis ( Ditlevsen, 1928) gen. et comb. nov., syn. nov.
= Diplopeltis asymmetricus Allgén, 1935 syn. nov. (note #4)
= Neodiplopeltula asymmetrica ( Allgén, 1935) gen. et comb. nov., syn. nov.
= Diplopeltis incisus sensu Gerlach, 1950 partim (note #1)
= Diplopeltula incisa sensu Voronov, 1982 partim (note #2)
= Diplopeltis ovalis sensu Sergeeva , 197 7 (note #5)
= Diplopeltula cuspidiboja Leduc, 2017 partim syn. nov. (note #6)
= Neodiplopeltula cuspidiboja ( Leduc, 2017) gen. et comb. nov., syn. nov.
Neodiplopeltula bathmanni ( Jensen, 1991) gen. et comb. nov.
= Diplopeltis bathmanni Jensen, 1991
= Diplopeltula bathmanni ( Jensen, 1991)
= Diplopeltula incisa sensu Vitiello, 1972 (note #7)
= Diplopeltula incisa sensu Voronov, 1982 partim (note #2)
= Diplopeltula cuspidiboja Leduc, 2017 partim syn. nov. (note #8)
= Neodiplopeltula cuspidiboja ( Leduc, 2017) gen. et comb. nov., syn. nov.
Neodiplopeltula indica (Gerlach, 1962) gen. et comb. nov. (note #9) = Diplopeltula indica Gerlach, 1962
Neodiplopeltula intermedia (Gerlach, 1954) gen. et comb. nov. (note #9) = Diplopeltula intermedia Gerlach, 1954
Neodiplopeltula obesa (Nguyen Vu Thahn, Nguyen Thahn Hien & Gagarin, 2012) gen. et comb. nov. = Diplopeltula obesa Nguyen Vu Thahn, Nguyen Thahn Hien & Gagarin, 2012 (note #9)
Neodiplopeltula onusta ( Wieser, 1956) gen. et comb. nov.
= Diplopeltis onustus Wieser, 1956
= Diplopeltula onusta ( Wieser, 1956)
= Diplopeltula incisa sensu Voronov, 1982 partim (note #2)
= Diplopeltula tchesunovi Fadeeva & Mordukhovich, 2013 syn. nov. (note #10)
= Neodiplopeltula tchesunovi ( Fadeeva & Mordukhovich, 2013) gen. et comb. nov., syn. nov.
Species inquirenda et incertae sedis
Diplopeltula breviceps Gerlach, 1950
Diplopeltula longiceps Gerlach, 1950
Species incertae sedis
Diplopeltula striolata Vincx & Gourbault, 1992 (note #11)
Diplopeltula ostrita Boucher & Helléouët, 1977 (note #12)
Nomenclatorial changes and notes
1) The description of Diplopeltis incisus sensu Gerlach, 1950 is based on specimens collected in several localities in the Kiel Bay and is likely to be a mixture of at least two different species: based on the oval shape and relatively small size of the periamphideal refractive plate, the position of cephalic setae posterior to the oral opening, and the position of the oral opening close to the anterior end, N. barentsi gen. et comb. nov. can be identified on fig. 3h, 3k & 3m in Gerlach (1950), while fig. 3b likely depicts N. incisa sensu stricto, based on the irregular shape and relatively large size of the periamphideal refractive plate, the position of cephalic setae posterior to the oral opening, and the position of the oral opening close to the anterior end. Unfortunately, exact measurements of morphological characters cannot be attained from the figures in Gerlach’s publication due to the absence of proper scale bars. Thus, our hypothesis on the taxonomic identity of specimens studied by Gerlach (1950) must solely rely on illustrations and cannot be verified any further.
2) The description of Diplopeltula incisa sensu Voronov, 1982 is based on specimens collected in six different locations from the White Sea, within an area of more than 100 × 200 km in size and ranging in depth from 50 to 250 meters. Many measurements given by Voronov vary within 2×–4× range. Morphological features of the anterior end also show a considerable variability (noted by Voronov himself) suggesting that his description is based on more than one morphospecies. Here we suggest that there are four different species of Neodiplopeltula gen. nov. illustrated in Voronov’s publication: fig. 1a likely depicts N. bathmanni gen. et comb. nov. with very narrow periamphideal plates and an oral opening located at level with the cephalic setae bases; both specimens depicted on fig. 1б and 1в match our concept of N. barentsi gen. et comb. nov. with relatively broad oval periamphideal plates and an oral opening located close to the anterior terminus, anterior to the cephalic setae bases; the presence of irregularly shaped periamphideal plates and an oral opening located close to the anterior terminus, anterior to cephalic setae bases in specimen depicted on 1г suggest it to belong to N. incisa gen. et comb. nov.; while the anterior end shown on fig. 1д completely matches with N. onusta gen. et comb. nov. (and confirmed by Voronov in his figure caption) in having periamphideal plates merging on the dorsal and ventral body sides and an oral opening located at level with the cephalic setae bases.
3) The holotype of Diplopeltis ovalis described by Ditlevsen (1928) from Greenland is available in the collection of the Natural History Museum of Denmark in Copenhagen and is in rather acceptable shape. Some morphological features, such as cephalic setae and the spinneret cannot be seen due to deterioration, while the vagina and rectum are difficult to observe since the specimen is not in perfect lateral position on the slide. Morphological and morphometric features that are visible and measurable, show no differences from N. barentsi gen. et comb. nov. – in particular, the position of the oral opening and morphology of refractive plates underlying the amphids are identical ( Figs 14A View Fig , 15 View Fig A–B). Consequently, Diplopeltis ovalis is being transferred to the genus Neodiplopeltula gen. nov. and synonimised with N. barentsi gen. et comb. nov.
4) The lectotype of Diplopeltis asymmetricus described by Allgén (1935) from Öresund was found in the collection and is in poor shape. The specimen is partly dried, thus making it impossible to observe and measure smaller features such as cephalic setae, spinneret and rectum. It is also slightly flattened, which affects a- and c’-ratios. Morphological features that are visible, show no differences from N. barentsi gen. et comb. nov. – in particular, the position of the oral opening and the morphology of refractive plates underlying the amphids are identical ( Figs 14B View Fig , 15C View Fig ). Consequently, Diplopeltis asymmetricus is being transferred to the genus Neodiplopeltula gen. nov. and synonimised with N. barentsi gen. et comb. nov.
5) Diplopeltis ovalis sensu Sergeeva, 1977 is similar to N. barentsi gen. et comb. nov. in the position of the oral opening on the dorsal side of the body but close to the anterior end, length and arrangement of the cephalic setae ( 7.5 µm vs 3.5–7.5 µm in present specimens), and the shape of refractive plates (elongated ovoid, not connected).
6) The male of Diplopeltula cuspidiboja gen. et comb. nov. is identical to N. barentsi gen. et comb. nov. in the position of the oral opening, size and shape of refractive plates and the great majority of quantitative characters (see Table 3 View Table 3 ), except for a slightly longer amphid ( 32 µm vs 21–25.5 µm) and gubernaculum ( 20 µm vs 10–11.5 µm).
7) Diplopeltula incisa sensu Vitiello, 1972 is similar to D. bathmanni in the position of the oral opening, length and the arrangement of the cephalic setae, the size and position of the periamphideal refractive plate, length of the tail and position of the caudal gland openings.
8) The female of Diplopeltula cuspidiboja gen. et comb. nov. is identical to N. bathmanni gen. et comb. nov. in the position of the oral opening, the position of the cephalic sensilla, the size and shape of the refractive plates ( Leduc 2017); as a result of having a larger body ( 2076 µm vs 1270–1673 µm), many morphometric characters of the female of D. cuspidiboja are also outside
the measurement range of both Nordic populations, which can be attributed to geographic variability.
9) Diplopeltula indica and Diplopeltula intermedia both match the diagnostic characters of the genus Neodiplopeltula gen. nov.: amphids in the shape of an elongated loop, a well-developed subcylindrical stoma and outstretched ovaries. Diplopeltula obesa also has amphids in the shape of an elongated loop and outstretched ovaries, however, the structure of the stoma in this species is described as “not expressed”, leaving some doubts as to its exact morphology.
10) Diplopeltula tchesunovi is identical to D. onusta in all qualitative and quantitative characters ( Table 4 View Table 4 ), with the exception of longer refractive plates ( 41–48 µm vs 32–38 µm), which can be attributed to geographic variability.
11) Diplopeltula striolata is described based only on one male, thus the structure of the female reproductive system remains unknown, questioning its systematic affinities. It is similar to the genus Neodiplopeltula gen. nov. in the morphology of the amphid (in the shape of an elongated loop) and stoma (small but well developed, subcylindrical). The morphology of the pharynx (uniformly cylindrical) also matches that of Neodiplopeltula gen. nov. However, D. striolata strongly differs from Neodiplopeltula gen. nov. in having a strongly annulated cuticle, the position of the excretory pore at the level with the anterior part of the intestine, the presence of a spinneret, the absence of a gubernaculum. Although this species can easily be identified, and its validity is unquestionable, its taxonomic position remains unresolved as it cannot be assigned to any of the existing genera with confidence ( species incertae sedis).
12) All available type specimens of Diplopeltula ostrita were examined, but not all morphological characters can be observed. That includes the female reproductive system and the structure of the ovaries. However, D. ostrita strongly differs from Neodiplopeltula gen. nov. in having a strongly annulated cuticle, a minute buccal cavity, weak spicules and gubernaculum. Similar to Diplopletula striolata , D. ostrita is here considered to be a valid species, which, however, cannot be assigned to any existing genus with confidence and must be considered species incertae sedis.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Neodiplopeltula
| Holovachov, Oleksandr & Boström, Sven 2018 |
Neodiplopeltula incisa ( Southern, 1914 )
| Holovachov & Boström 2018 |
Diplopeltula incisa
| sensu Voronov 1982 |
Diplopeltis incisus
| sensu Gerlach 1950 |
Diplopeltis incisus
| Southern 1914 |