
Actinocyclus ferrarioae Lameiro, Vouilloud & Cefarelli, 2023
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.626.4.4 |
|
DOI |
https://doi.org/10.5281/zenodo.10249209 |
|
persistent identifier |
https://treatment.plazi.org/id/03ACBD51-FFA1-FFD6-FF72-CAC9FC76564C |
|
treatment provided by |
Plazi |
|
scientific name |
Actinocyclus ferrarioae Lameiro, Vouilloud & Cefarelli |
| status |
nom. nov. |
Actinocyclus ferrarioae Lameiro, Vouilloud & Cefarelli nom. nov.
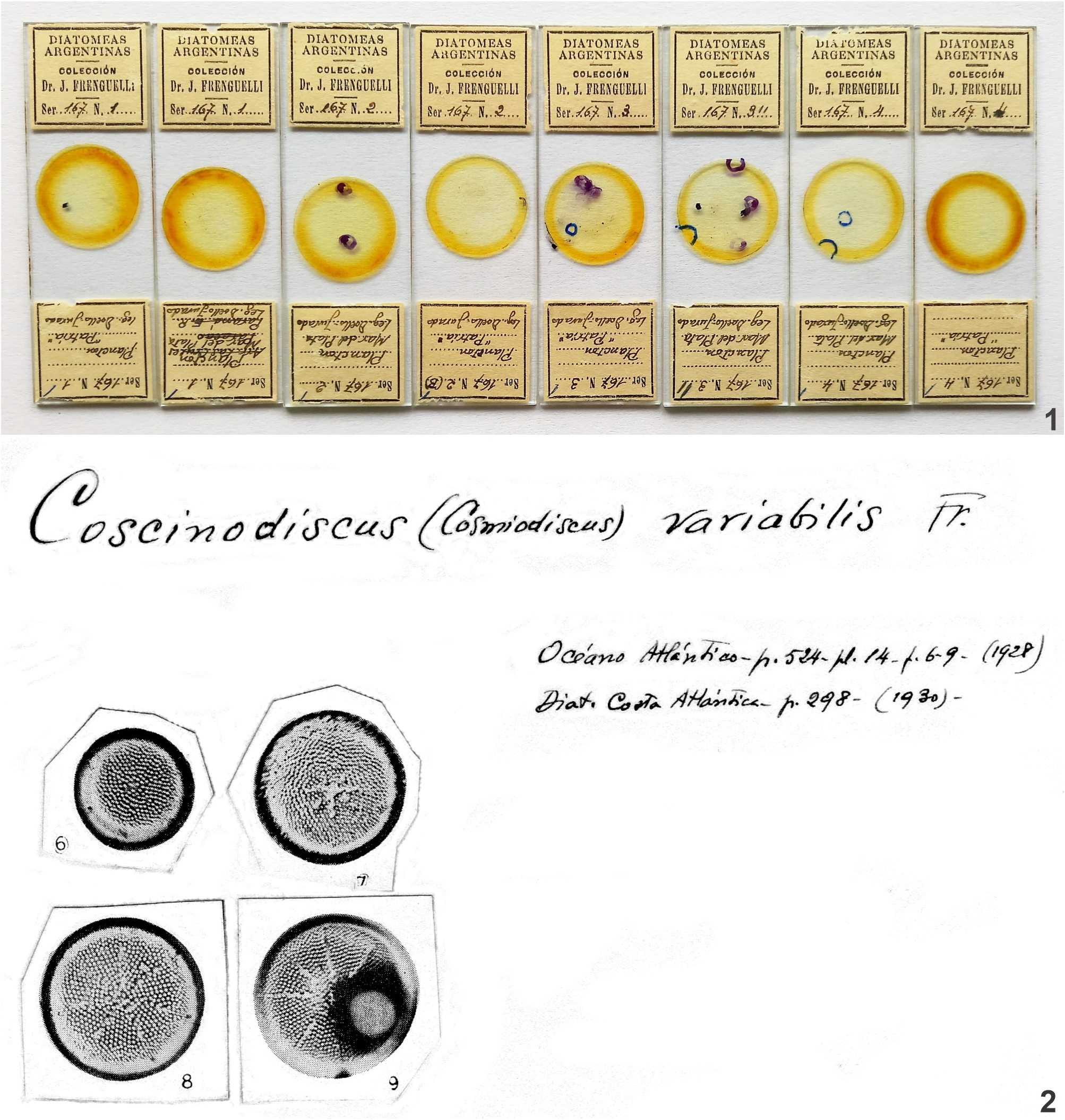
Replaced synonym: Coscinodiscus variabilis Frenguelli 1928 . Anales del Museo Nacional de Historia Natural “Bernardino Rivadavia”. Protistología 1: 524, pl. 14, figs 6–9.
Type: — ARGENTINA. Buenos Aires province, Mar del Plata, planktonic samples taken 5–30 miles from the coast of Mar del Plata, Atlantic Ocean , 38°S, 57°30´W. M. Doello-Jurado, February-April 1914. Lectotype (designated here):—ARGENTINA. Frenguelli Collection 167!, Specimen of pl. 14, fig. 8 of the original publication (See Fig. 2 View FIGURES 1–2 ). GoogleMaps
Hayashi et al. (2012) and Wu et al. (2022) analyzed the relation between the valve diameter and the number of rimoportulae and recognized that the ratio of number of rimoportulae and fascicles to valve size is an important criterion for the identification of polymorphic Actinocyclus species. We propose the RP density as a character to compare Actinocyclus species that are morphologically similar. RP density is a relatively stable character within each species and helped us identify Actinocyclus species remarking its usefulness as another diagnostic character for Actinocyclus taxa together with the fasciculation pattern or the striae or areolae density.
Hasle & Syvertsen (1997) proposed the existence of three different types of fasciculation in Actinocyclus species: 1) radial areolae parallel to a central row, 2) radial areolae parallel to a side row and 3) areolae rows parallel to a central and/or edge row. Actinocylus ferrarioae has rows of areolae parallel to a central row in each fascicle, a feature that remains constant in all specimens analyzed. It is also interesting to remark that on those specimens that lack evident hyaline rays, the fascicles are defined by conspicuous rows or areola that allow us to recognize its fasciculation pattern (figs 16,17, 22–25).
In other Actinocyclus species included in Table 1 View TABLE 1 , the fasciculation pattern shows some variation and even in some species this feature varies according to valve size as observed in A. nipponicus ( Hayashi et al. 2012) .
Given the great extent of morphological variation within Actinocyclus taxa regarding fasciculation, and structure of the hyaline rays, there are many fossil species morphologically similar that can be confused with A. ferrarioae under LM (Table I).
Our specimens with conspicuous and long hyaline rays resemble Actinocyclus asteriscus Wu et al. (2022: 3–4) but striae density is lower and RP density is higher in A. ferrarioae . Specimens of A. ferrarioae with incomplete hyaline rays inserting from the center of the valve face are morphologically similar to a group of Actinocyclus taxa including A. podolicus (Missuna) Koryzenko (1959: 33) only differing in the areolae density and A. nigriniae Barron (2005: 640) with whom varies in valve size and areola density. Specimens of A. ferrarioae with incomplete hyaline rays located in the middle area of the valve face and close to the valve margins are similar to A. kanitzii (Panctosek & Grunow) Shauderna (1983: 4–6) but have smaller valve sizes and a lower RP in 100 µm and A. ingens var. indica Desichakary & Prema (1987: 5) but have smaller valve sizes and higher areolae density. A. ingens var. indica is morphologically quite similar to some of our specimens but having only one specimen illustrated in the original description of the species does not allow us to make a complete comparison between the taxa. Specimens of A. ferrarioae lacking hyaline rays are similar to A. tenellus (Brebbison) Andrews (1976: 14) but have smaller valve sizes along with more RP in 100 µm and a much lower stria density and also resemble A. claviolus Bradbury & Krebs (1995: 5–6) which has a distinctive central annulus absent in A. ferrarioae which also has slightly lower stria density ( Table 1 View TABLE 1 ).
It is also interesting to analyze the extension of the hyaline rays to understand the morphological variability that undoubtedly characterizes Actinocyclus recent and fossil taxa. Wu et al. (2022) studied fossil Actinocyclus taxa from Oligocene marine deposits and proposed the existence of six groups of species (A–E) considering the hyaline rays structure. Following this characterization, some of our specimens would fit into Groups A and B, exhibiting large and well developed hyaline rays that cover the entire or nearly all the valve face, connecting both with the valve center and margins along with other specimens with shorter hyaline rays that reach the valve margins but only cover half of the valve face without reaching its center. Other specimens would fit into Group C with hyaline rays inserting from the center of the valve face and not reaching the valve margins along with other specimens with shorter hyaline rays that reach the valve margins but only cover half of the valve face without reaching its center. Finally, the specimens lacking hyaline rays would be included in between Groups E–F with a clear variation regarding valve size, definition of valve fascicles and their boundaries. Wu et al. (2022) proposed that larger specimens with conspicuous hyaline rays can be considered as basal forms and smaller ones with no conspicuous hyaline rays as modern forms, suggesting that the reduction of the hyaline rays is a result of environmental changes throughout the planet´s evolution which led to the diversification of the species in continental environments. The morphological variability within Actinocyclus taxa makes us wonder whether the evolution of the hyaline rays is directed to the reduction or the complexion of these structures. The presence of high variation in the development of hyaline rays in a recent species ( A. ferrarioae ) contrast with Wu et al. (2022) hypothesis of the evolution of this structure towards its reduction. However, there can be convergent evolution to generate similar morphologies, or perhaps the recent specimens evolved from extinct species of Actinocyclus (D.M. Harwood pers. comm.).
Regarding its distribution, A. ferrarioae was originally reported for the Atlantic coasts of Buenos Aires province and a few decades later Mueller-Melchers (1955) reported it for the Atlantic coasts of Uruguay and southern Brazil but he only mentioned the species and did not include any illustration of it. Krasske (1934) analyzed fossil material from Germany and identified it as Coscinodiscus variabilis Frenguelli but analyzing the photos included in that publication we concluded that this was a misidentification, given that our specimens of A. ferrarioae have a more thickened structure of the rimoportulae and a higher areolae density and valve size. From these data, we can consider that currently the geographic distribution of A. ferrarioae is in the South Atlantic Ocean between Argentina and Brazil. Actinocyclus ferrarioae was also registered by Frenguelli (1930) as C. variabilis near Mar del Plata City in material obtained from scraping of a marine gastropod, a register that was confirmed in the present study. This finding can be interpreted as a possible epibiosis case but could also be accidental due to possible contamination of the original material during sampling. The complete ecological characterization of the species would require current material in order to make a significant ecological description.
Finally, it seems important to make some remarks regarding the analysis of collection material. When analyzing this type of material, it is frequent to obtain data that differs with that given by the authors in their original publications. Discrepancies between original measurements by Frenguelli and those obtained when his materials are re-examined are rather usual ( Kociolek & Vouilloud 2020 and references therein) and, partially, can be related with the lower number of specimens analyzed by Frenguelli, a frequent practice at that time. This demonstrates the importance of reviewing the original materials rather than relying on previous reports. Future studies regarding current materials from the same areas would help complete the characterization of A. ferrarioae under SEM.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Actinocyclus ferrarioae Lameiro, Vouilloud & Cefarelli
| Lameiro, Rubén A., Cefarelli, Adrián O. & Vouilloud, Amelia A. 2023 |
Coscinodiscus variabilis
| Frenguelli 1928 |