
Macrophthalmus (Mareotis) pistrosinus, Davie, Peter J. F., 2008
|
publication ID |
https://doi.org/ 10.5281/zenodo.182729 |
|
DOI |
https://doi.org/10.5281/zenodo.6231930 |
|
persistent identifier |
https://treatment.plazi.org/id/03AF5539-FF80-FFD5-FF5C-70DC7FCC4D11 |
|
treatment provided by |
Plazi |
|
scientific name |
Macrophthalmus (Mareotis) pistrosinus |
| status |
sp. nov. |
Macrophthalmus (Mareotis) pistrosinus View in CoL sp. nov.
( Figs. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Macrophthalmus (Mareotis) japonicus Barnes, 1967: 224 View in CoL –226, fig. 8, pl. 2(d). Macrophthalmus (Mareotis) aff. japonicus Kitaura et al., 2006: 46 View in CoL .
Material Examined. Holotype: WAM 655-65, 1 male (30.9 x 20.4 mm), Teggs Channel, Shark Bay, WA, 14.8.1963, R. Slack-Smith. Paratypes: WAM 655-65, 2 males (26.6 x 14.1, 22.0 x 15.1 mm), same data as holotype above; WAM 651-65, 1 male, Denham Hummock, Shark Bay, WA from burrow near mangrove creek, 9.1.1963, R. Slack-Smith.
Description. Front deflexed, constricted between bases of ocular peduncles; margin smooth in part but inner portion covered with small, tubercular granules, with granular surface, bilobed anterior margin, deep median furrow.
Upper orbital border with slightly oblique, sinuous margin with large tubercular granules along whole length. Lower orbital border similarly studded with large tubercular granules; inner four-fifths straight, outer fifth abruptly sloping.
Carapace with 2 large and 1 small anterolateral teeth. External orbital angle pronounced, large, broad, rectangular, pointed anteriorly, directed outwards, slightly forwards; separated from second lateral tooth by wide U-shaped incision; outer margin with large tubercular granules; anterior margin with granules continuous with those of upper orbital border. Second lateral tooth large, broad, somewhat triangular, directed outwards and projecting beyond external orbital angle; separated from third lateral tooth by small but distinct incision; both margins with tubercular granules, outer margin slightly convex. Third lateral tooth small, conical, projecting outwards; outer margin with tubercular granules.
Carapace surface, excepting small central area, entirely covered by large granules; with deep, wide, setaecontaining furrows clearly demarcating regions; with transverse granular and setose row extending across anterior branchial region from level of third lateral tooth, with similar transverse row above insertion of fourth pereiopod, and with 2 longitudinal rows of granules and setae branchially, subparallel to each other and to posterolateral carapace margins. Greatest carapace breadth across second lateral teeth, behind which lateral margins subparallel. Lateral margins with large granules, row of setae.
Ocular peduncles long, narrow; cornea extending almost to tip of external orbital angle.
Central region of epistome distinctly concave.
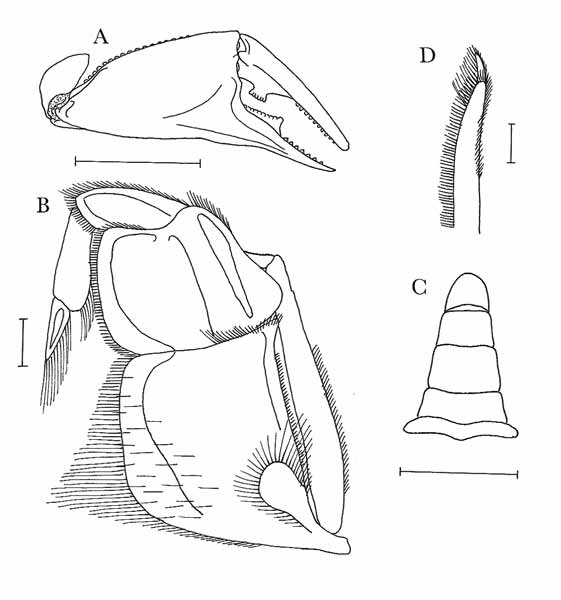
Male cheliped (see Figs. 1 View FIGURE 1 A & 3). Merus elongated, trigonal. Inner margin setose; upper margin with distal row of tubercular granules; outer margin finely granular. Outer surface with few scattered granules; inner and lower surfaces heavily setose, with scattered granules over remainder. Carpus almost glabrous, with only patches of fine setae on lower half. Upper margin with row of large tubercles; lower margin finely granular. Outer surface granular towards margins, more or less smooth centrally; inner surface with serrated crest of large tubercular granules, highest in centre. Palm (= manus) elongated. Upper margin with longitudinal row of large tubercles; lower margin finely granular. Outer surface finely granular, granules increasing in size towards carpus, without longitudinal ridge near lower margin, with slight depression near base of index; inner surface granular, with narrow longitudinal band of setae along uppermost third. Index (= pollex, immovable finger) deflexed, glabrous. Outer surface granular near upper and lower margins; inner surface granular. Lower margin feebly granular proximally, smooth distally; cutting margin with large, wedge-shaped, crenulated tooth occupying proximal half, distally with few large and many small granules. Dactylus curved, glabrous. Outer surface with granules near upper and lower margins; inner surface granular. Upper margin granular; cutting margin with large, quadrangular, crenulated tooth proximal to margin’s centre, wider at tip than at base, with distal row of tubercular granules.
Pereiopods. Meri elongated; upper, lower margins parallel, with rows of large granules; lower surfaces granular. Carpi granular, with distinct longitudinal ridges. Propodi granular, with faint longitudinal ridges. Dactyli lanceolated.
Male abdomen (see Fig. 1 View FIGURE 1 C). Lateral margins of segments 4–6 almost straight; lower margins of telson and 6th segment almost straight, lower margins of 5th and 4th segments concave. Proximal and posterior margins of sternal segments granular.
External (third) maxilliped (see Fig. 1 View FIGURE 1 B) with merus markedly smaller than ischium, with raised areas of merus, ischium pitted. Internal, external margins of ischium slightly concave to almost straight, subparallel. Internal margin of merus slightly convex; external margin with marked posteroexternal convexity.
First male pleopod ( Fig. 1 View FIGURE 1 D) slightly curved, with marked torsion, with well-developed terminal lobe and setae on internal margin distally.
Dimensions. Maximum known size: 30.9mm carapace breadth. Breadth of front from 15% of carapace width at 10mm carapace breadth to <10% at 30mm carapace breadth. Carapace length 67–70% carapace breadth.
Distribution and habitat. The species is known in Shark Bay from Carnarvon (24.9 0S, 113.7 0E) to Long Point (25.5 0S, 113.9 0E), along the north-eastern shore, in burrows in well-drained sandy and muddy sediments adjacent to both salt-marsh and mangroves, habitats essentially similar to those of M. japonicus and M. banzai , although perhaps M. pistrosinus sp. nov. occurs at higher tidal levels than those species (see Kitaura et al. 2006).
Etymology. Named after its single known locality, Shark Bay (Latin: sinus ‘a marine bay or gulf’; pistris or pistrix ‘a sea monster, such as a shark, saw-fish or whale’). The name is used as a noun in apposition.
Remarks. Morphologically, the species is extremely close to M. japonicus and its waving display is also similar, but it differs behaviourally by showing burrow-plugging (see below) which does not occur in either of the northern M. japonicus -complex species ( Kitaura et al. 2002) The following morphological characters are also useful for distinguishing the three species within the complex:
1. On M. pistrosinus sp. nov. there is a dense longitudinal band of fine setae on the inner surface of the palm of the male chela, whereas in M. japonicus and M. banzai the inner surface is glabrous, except in M. japonicus for a single row of fine setae lying close to the upper margin (this is a character present on the Queensland Museum sub-adult male specimen, but which appears not to have been mentioned in other accounts of this species).
2. The abdomen of male M. pistrosinus sp. nov. is relatively narrower than that of M. japonicus . In particular, the 5th and 6th segments and telson differ in the following breadth/length proportions ( M. pistrosinus sp. nov. first): 1.84 vs. 2.40; 1.94 vs. 2.18; 1.32 vs. 1.47. The proportions for M. japonicus are based on the single male in the QM collection, and from the figures of Shen (1932: fig. 134a), Kim (1973) and Wada & Sakai (1989); all of which agree closely in overall appearance and segment proportions.
3. The external orbital angle of M. pistrosinus sp. nov. is more pointed laterally than in M. japonicus - c.f. Figs. 2 View FIGURE 2 & 4 View FIGURE 4 , Shen (1932: fig. 132).
4. The upper and lower margins of the meri of the walking legs are nearly parallel in M. pistrosinus sp. nov. whereas they are gently convex in M. japonicus .
Barnes (1967) remarked that his Australian specimens, here described as M. pistrosinus sp. nov., appeared to differ from typical M. japonicus in the form of the longitudinal granular ridges on the branchial regions of the carapace. These ‘atypical’ ridges were later shown also to occur in material of M. japonicus from China ( Barnes 1970).
The burrow [construction] and social behaviour of the new species (waving display, fighting behaviour and courtship) have been documented by Kitaura et al. (2002). The chelipeds are moved vertically during waving displays, as in M. japonicus , although the duration of the interval from the apex position to the final lowered one was significantly longer in M. pistrosinus . During fighting behaviour, mutual pushing of the chelipeds occurs as the chelipeds cross over during lateral lowering (“stage B”), a stage not seen in M. japonicus or M. banzai . The new species also differs from the two latter ones in closing its burrow with a plug of mud collected near the burrow entrance, as also seen in fiddler crabs ( Uca ) and several dotillids.



| WAM |
Western Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Macrophthalmus (Mareotis) pistrosinus
| Davie, Peter J. F. 2008 |
Macrophthalmus (Mareotis) japonicus
| Kitaura 2006: 46 |
| Barnes 1967: 224 |