
Calyptocephalella sp., 2020
|
publication ID |
https://doi.org/10.1038/s41598-020-61973-5 |
|
DOI |
https://doi.org/10.5281/zenodo.3803362 |
|
persistent identifier |
https://treatment.plazi.org/id/03AF87E1-EC5E-FFFF-FE52-F92A6171F9E8 |
|
treatment provided by |
Plazi |
|
scientific name |
Calyptocephalella sp. |
| status |
|
Calyptocephalella sp.
Figures 2 View Figure 2 and 3 View Figure 3
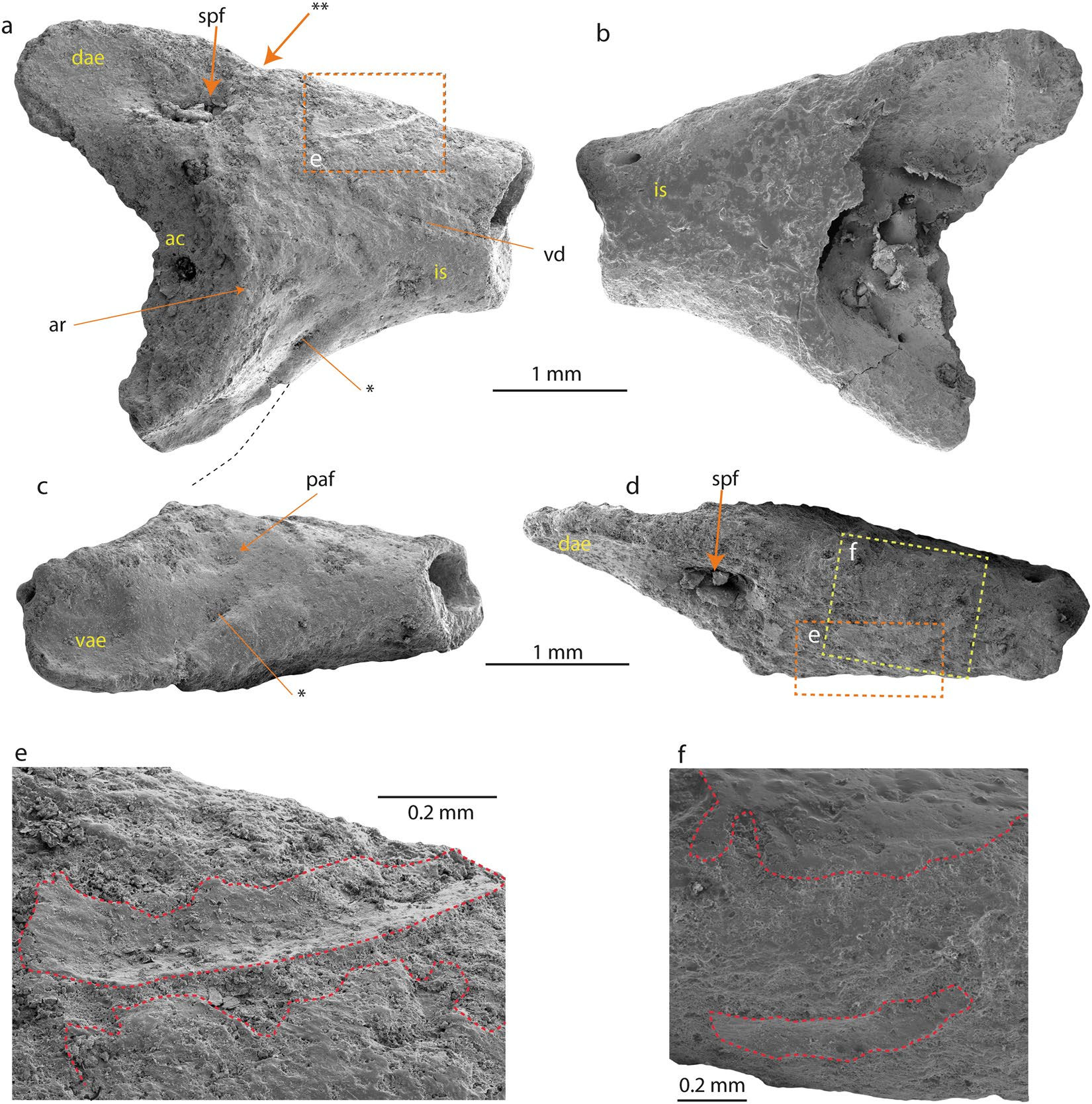
Referred specimens. Swedish Museum of Natural History NRM-PZ B282 , right ilium ( Fig. 2 View Figure 2 ), NRM-PZ B281 , skull bone ( Fig. 3 View Figure 3 ). GoogleMaps
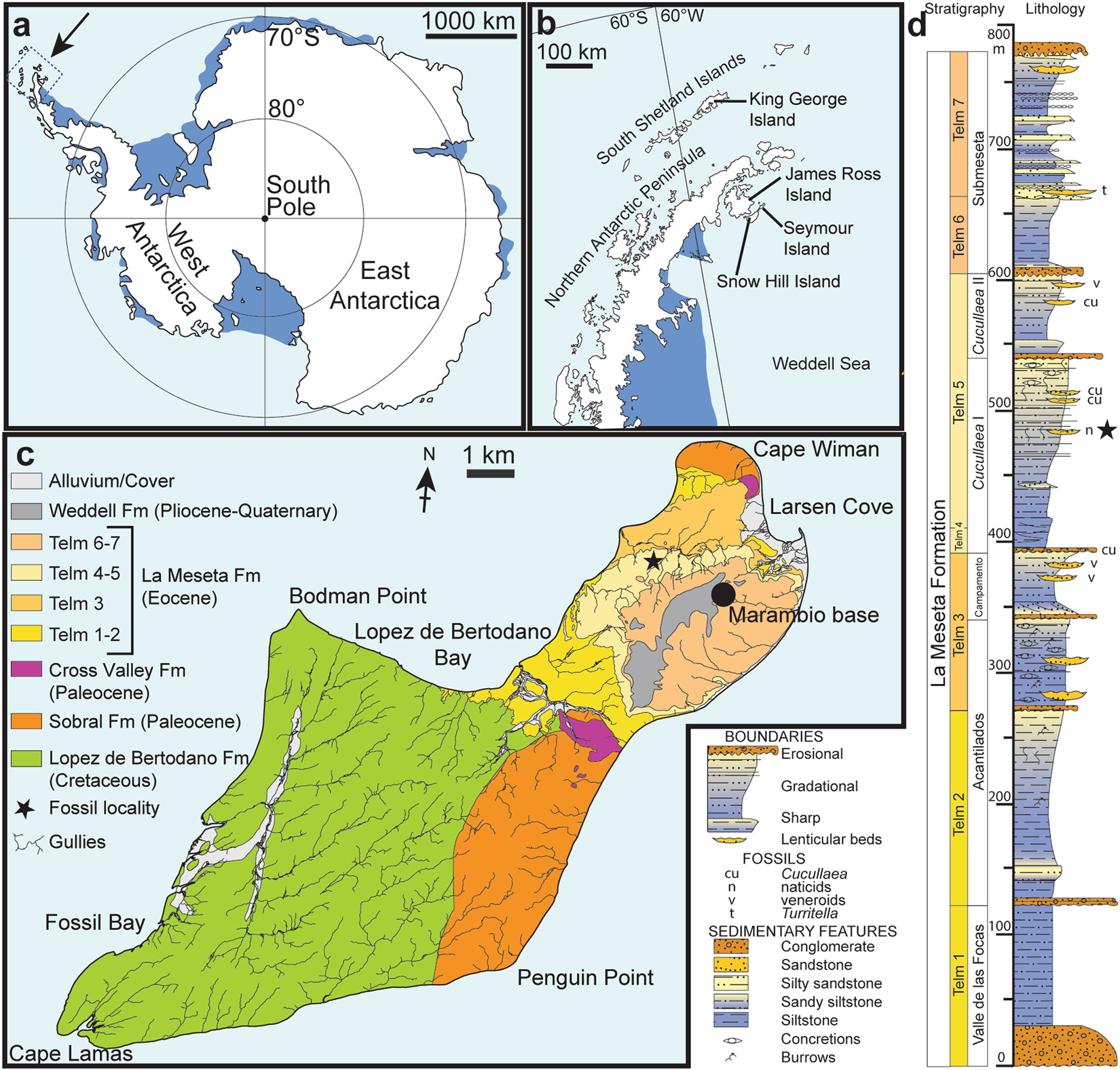
Locality, horizon and age. IAA 2/95, Marsupial site, Seymour Island, Antarctic Peninsula GoogleMaps ( 64°13′58″S; 56°39′06″W). ‘ Natica horizon’ within the Cucullaea I Allomember (Telm 5) of the La Meseta Formation, Bartonian (40Ma), Eocene GoogleMaps 23, 24 ( Fig. 1 View Figure 1 ).
Measurements. The preserved part of the ilium measures 3.9 mm in length, the distance from the tip of the dorsal acetabular expansion to the (preserved) tip of the ventral acetabular expansion measures 3.3mm, the highest height of the acetabular fossa equals 2.5 mm. The skull bone measures 2.7 mm at its both broadest and longest parts.
Ilium. The fragmentary right ilium (NRM-PZ B282) lacks the caudal portion of the acetabulum and most of the iliac shaft. The dorsal acetabular expansion has a smooth lateral surface and is higher than the preserved part of the ventral acetabular expansion ( Fig. 2a View Figure 2 ). A large and deep supraacetabular fossa is present at its base ( Fig. 2a,d View Figure 2 ). The preserved portion of the acetabulum is concave and its shape allows concluding a (semi-)circular outline. The acetabular rim is most prominent at its anterior part ( Fig. 2a,c View Figure 2 ). The barely-developed ventral acetabular expansion projects ventrally. The posterior-most portion of the ventral acetabular expansion is broken off. However, the anterior portion of the ventral acetabular expansion is higher than the preserved posterior portion. In ventral view ( Fig. 2a,c View Figure 2 ), the lateral surface of the ventral acetabular expansion is convex. The ventral acetabular expansion possesses a shallow and broad depression. In the preacetabular zone, a small and shallow preacetabular fossa is present ( Fig. 2c View Figure 2 ). The preserved portion of the iliac shaft is damaged and precludes a confident statement whether the dorsal protuberance is present or absent. A narrow and shallow longitudinal groove is observable in the lateral surface of the iliac shaft, which probably corresponds to the posterior extension of the ventral depression (sensu 10) ( Fig. 2a View Figure 2 ). However, intact parts of bone surface are preserved slightly ventral to the dorsal margin on both lateral and medial surfaces ( Fig. 2e,f View Figure 2 ). The one on the lateral surface is a curved shallow groove and runs posteroventrally ( Fig. 2e View Figure 2 ). This feature anteroventrally demarcates the slightly elevated roughened scar interpreted above as the dorsal protuberance. The area between the dorsal acetabular expansion and iliac shaft is slightly projected dorsally. This area corresponds to the position of the dorsal protuberance. In fact, no clear evidence of a dorsal protuberance can be found on the ilium, only a slightly roughened area with minimal elevation that corresponds to the dorsal protuberance and the scar for the insertion of the musculus gluteus magnus can be observed. At the caudal side of the dorsal protuberance, a distinct notch is visible ( Fig. 2a View Figure 2 ) which we consider as a further evidence of our interpretation. The area corresponding to the dorsal protuberance is located anteriorly to the anterior margin of the acetabular rim. Medially, the entire surface opposing the acetabulum is lost and the area preserved more anteriorly is slightly convex medially and smooth. Anteriorly and dorsally, just adjacent to the anterior end of the dorsal protuberance a foramen is present ( Fig. 2b View Figure 2 ).
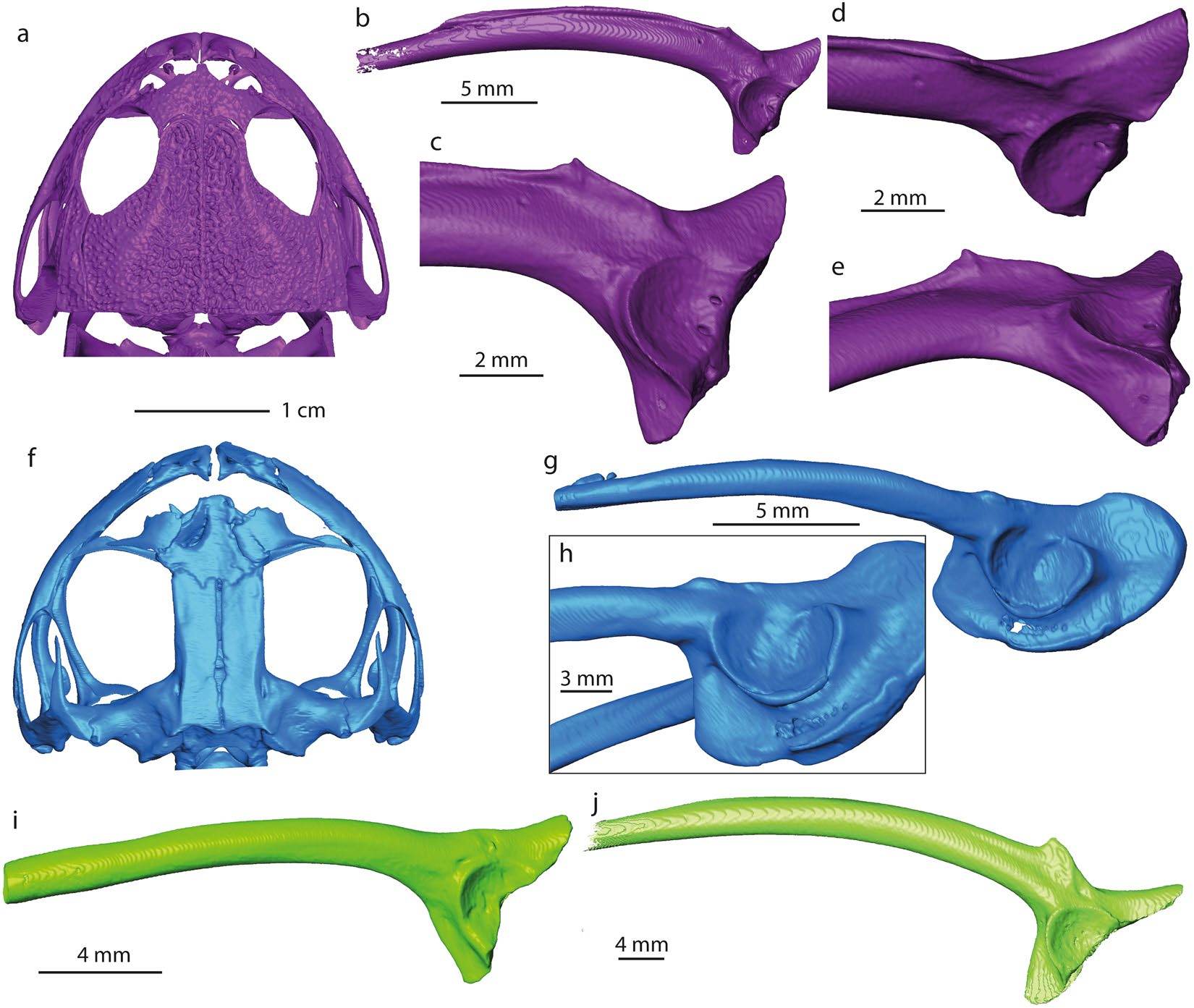
The fragmentary right ilium can be referred to an anuran based on the following characters 30 (the numbers before the characters correspond to the feature numbers of Appendix 1 in Gardner et al. 30): 7. (semi-) circular acetabulum; 9. acetabulum with distinct margins; 10. acetabular surface concave; 13. at least dorsal acetabular expansion is strongly divergent; 18. the dorsal protuberance present. Thus, the ilium derives from a small-sized frog (3.8 ± 0.4 cm snout-vent length, see methods, Table 1). The specimen is partly eroded and rather poorly preserved; however, it can be compared with all South American and Australian frog families ( Figs. 4 View Figure 4 , S 1 View Figure 1 and S 2 View Figure 2 , Table S1). The families Ranidae , Bufonidae and Hylidae have not been illustrated in the present work, since their morphology is well known 31 ( Table S1). The comparison has been done at family level, since the ilia display diagnostic features characteristic for identification of the family (dimensions of the dorsal and ventral acetabular expansions; location of the dorsal protuberance relative to the anterior margin of the acetabular rim etc. 32, 33). The studied ilium (NRM-PZ B282) differs in: (1) Reduced anterior portion of the dorsal acetabular expansion from nearly all South American and Australian frog families and the genus Telmatobufo , which have moderately or strongly developed anterior portion of the dorsal acetabular expansion. Only the genus Calyptocephalella ( Fig. 4c,e View Figure 4 ), the families Ranidae 31, Pipidae ( Fig. S1a View Figure 1 ), Rhinodermatidae ( Fig. S1j View Figure 1 ), and Leptodactylidae ( Fig. S2b View Figure 2 ) have similar state/morphology of this character. (2) Dorsal protuberance located either at the level of or anteriorly from the anterior margin of the acetabular rim from nearly all families, besides Brachycephalidae ( Fig. S1a View Figure 1 ), Rhinodermatidae ( Fig. S1j View Figure 1 ), Telmatobiidae ( Fig. S1k View Figure 1 ), Hyloididae ( Fig. S1m View Figure 1 ), Leptodactylidae ( Fig. S2b View Figure 2 ) and the genera Calyptocephalella ( Fig. 4 View Figure 4 b–e) and Telmatobufo ( Fig. 4g,h View Figure 4 ). (3) Developed dorsal acetabular expansion from the families Ranidae 31, Hylidae 31, Bufonidae 31, Myobatrachidae ( Fig. 4i View Figure 4 ), Pipidae ( Fig. S1i View Figure 1 ), Microhylidae ( Fig. S1b View Figure 1 ), Telmatobiidae ( Fig. S1k View Figure 1 ), Leptodactylidae ( Fig. S2b View Figure 2 ), Allophrynidae ( Fig. S2c View Figure 2 ), Centrolenidae ( Fig. S2d View Figure 2 ) and the genus Telmatobufo ( Fig. 4g,h View Figure 4 ). Other families have moderately or well-developed dorsal acetabular expansion, however, due to incomplete preservation of the Antarctic frog remain any further comparison is impossible. (4) Weakly developed dorsal protuberance and lack of dorsal tubercle from nearly all families (e.g. Limnodynastidae , Fig. 4j View Figure 4 ), besides Calyptocephalellidae ( Fig. 4 View Figure 4 b–e,g,h), Myobatrachidae ( Fig. 4i View Figure 4 ), Craugastoridae ( Fig. S1e View Figure 1 ), and Dendrabatidae ( Fig. S2e View Figure 2 ).
Among the compared forms, only the South American endemic genus Calyptocephalella resembles all mentioned four characters. In addition to this, a shallow and broad depression on the anterior portion of the ventral acetabular expansion is a unique character observable on our ilium (NRM-PZ B282) and Recent Calyptocephalella gayi ( Fig. 4 View Figure 4 ). Further, the fossil ilium displays a ventral depression on its lateral surface anteriorly to the acetabulum ( Fig. 2c View Figure 2 ), a comparable structure can be observed also in the fossil species Calyptocephalella canqueli 9 but not in the Recent species C. gayi ( Fig. 4 View Figure 4 ).
Skull element. The second bone fragment (NRM-PZ B281) is flat and slightly curved. Both sides of the bone have different structures. One surface is covered by small to large circular or reniform in outline, rather deep pits, which sink in the planar surface of the bone ( Fig. 3a View Figure 3 ). The diameters of pits vary from 0.1–0.7 mm and some of them are punctured by foramina. The opposite surface of the bone is in general smooth, slightly deepened and is pierced with some foramina, some of which are preceded by a groove ( Fig. 3b View Figure 3 ). One side of the fragment preserves an unbroken margin of the original bone with a distinct process that is bent and that gives the bone a curved shape ( Fig. 3c View Figure 3 ). The ornamented surface of the bone projects slightly over this process. Comparable ornamentation, build of pits of different size, is found on the dorsal surfaces of different cranial and postcranial bones of amphibians and reptiles 34, 35. Among them, the following groups can be excluded from consideration: (1) Albanerpetontidae (Allocaudata) ; albanerpetontids are a primary Laurasian lissamphibian group with a single occurrence in Northern Africa. So far no evidence of a Gondwanan radiation of albanerpetontids exists 36. In addition to this, all their ornamented bones (e.g. frontal, premaxillae) 37 do not resemble the bone described here. (2) Caudata ; salamanders are also considered as a Laurasian group, with a number of occurrences in Africa which need critical revision 38. In salamanders, ornamented bones are found both among skull bones and on vertebrae (on plates located on the tip of the neural arch) 39. Bone ornamentation here (e.g. Tylototriton, Chelotriton, Echinotriton 39, 40) is represented by a network of pits, ridges and pointy spines that do not resemble the bone described here. (3) Crocodylia; in crocodyliforms, comparable patterns of ornamentation with well-developed pits appear only with growth during later ontogenetic stages 41 – 43. On one hand, the bone dimensions indicate a small-sized animal (corresponding to a juvenile crocodilian without such developed ornamentation). On the other hand, crocodylian osteoderms are flat without any processes, unlike the studied bone. (4) Testudines ; shell plates of several turtles, such as Trionyx, Allaeochelys etc. 35, are also covered by ornamentation. The ornamentation is characterized by larger and closely arranged pits, which are not always clearly delimited from each other (see Scheyer 35: Fig. 1a View Figure 1 ). (5) Lacertilia; lizards also have skull bones and osteoderms covered with ornamentation patterns 44. They all are characterized by a network of spines, grooves, ridges 45 and protuberances 46, which differs from the morphology on NRM-PZ B281.
The ornamentation pattern found in NRM-PZ B281 is comparable to that of some frog genera, i.e. Thaumastosaurus 47, Beelzebufo 48, Calyptocephalella 9 and Baurubatrachus 49, but only the last three genera are Gondwanan forms and, thus, considered for comparison herein. Beelzebufo is a very large form and the ornamentation pattern is present both on skull bones and on vertebrae 50. Calyptocephalella 9 and Baurubatrachus 49 have very similar ornamentation patterns on the surfaces of hyperossified skull bones, comparable to our specimen. A recent phylogenetic analysis 49 placed the Late Cretaceous Baurubatrachus within both Recent calyptocephalellid genera Calyptocephalella and Telmatobufo . Though Muzzopappa and Báez 10 mention that both Calyptocephalella and Telmatobufo are characterized by a heavily ossified neurocranium, we can confirm this only for the former genus ( Fig. 4a,f View Figure 4 ). Within Calyptocephalella the ornamentation pattern on skull bones is variable. In C. conquella 10, it is built either by network of pits in small individuals, or tuberculated ornamentation in adults. In C. satan 9 and C. casamayorensis 51, ornamented skull bones are slightly larger than NRM-PZ B281 but they have a similar pattern built of pits. C. pichifleufensis 48 is known by larger individuals which show similar ornamentation patterns but with larger pits. In comparison to these species, the Antarctic frog displays an ornamentation most similar to that of C. satan 9 and C. casamayorensis 48. Taking into account our comparison, we conclude that the ornamented bone fragment NRM-PZ B281 represents a skull bone (most probably a nasal) of a small-sized Calyptocephalella or Baurubatrachus . Given the presence of a small Calyptocephalella as indicated by the ilium in the same, only few m 2 measuring outcrop, it is most likely that specimen NRM-PZ B281 belongs to the same genus. A comparable record of an ilium and ornamented bones referable to the genus Calyptocephalella has been mentioned in Báez 52.
Discussion
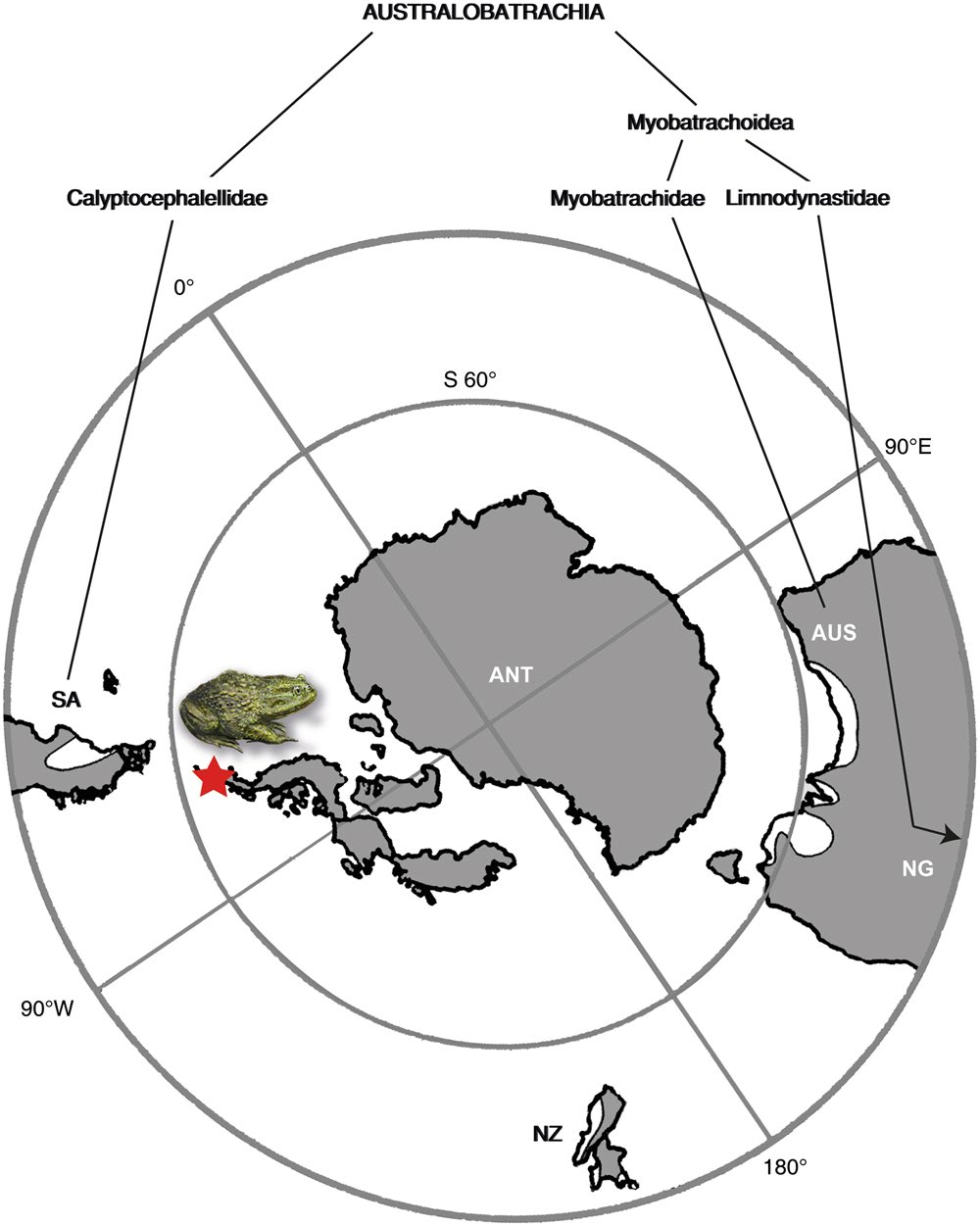
Among Recent amphibians, the frogs ( Anura ) have the widest distribution, covering all continents except Antarctica, where the conditions have been uninhabitable for over tens of millions of years. Contrary to all other continents, no traces of any extant amphibian group, all of which belong to the lissamphibian clade, have been documented from Antarctica. This paper presents the first record of a lissamphibian in Antarctica, with Eocene fossils referable to the order Anura , and most likely to the australobatrachian genus Calyptocephalella . The family Calyptocephalellidae belongs to neobatrachian frogs and is exclusively known from South America 53, 54. The five extant species, including the monospecific genus Calyptocephalella with hyperossified skull bones, are restricted to the Chilenean Andes 54 while most fossil representatives are known from Argentine Patagonia 9, 48, 53. Today, Calyptocephalella inhabits lowland areas of central Chile (upper elevation limit 500 m) east of the Andes within temperate and humid climates, between latitudes 30–43°S. It has an aquatic or semiaquatic lifestyle and populates standing or slow flowing water bodies (lakes, ponds, streams) in the Valdivian temperate Nothofagus forests 54 – 56.
The oldest fossils referable to Calyptocephalella are known from the Upper Cretaceous of Argentina 9, 52. During the Paleocene–terminal early Miocene, their geographic range was restricted to Patagonia east of the Andes 48, 51, 53, 57. Not until the late Pleistocene did they appear west of the Andes, where they have their endemic present-day distribution 54 – 57.
The clade Australobatrachia comprises Myobatrachoidea (families Myobatrachidae + Limnodynastidae sensu 27), nowadays distributed in Australia and south of New Guinea, and the family Calyptocephalellidae ( Batrachophrynidae sensu 27). Australobatrachia are considered as a stem group of the Hyloidea. The earliest myobatrachoid from Australia is at least as old as early Eocene, based on fragmentary ilia that were referred to the basal extant Lechriodus 58. The split between Calyptocephalellidae and Myobatrachidae ( Calyptocephalellidae + Myobatrachoidea sensu 27) occurred ~100 Ma (~Early-Late Cretaceous boundary) 59. Considering the distributions of extant Australobatrachia ( Fig. 5 View Figure 5 ), the earliest fossil records 10 and the divergence age (from genetic data) 59 of both Calyptocephalellidae and Myobatrachoidea lineages, it is clear that Antarctica had played an important palaeobiogeographic role for Australobatrachia and their consequent dispersal. Because the most recent common ancestor of the clade, including Hyloidae and Myobatrachidae + Calyptocephalellidae , occurred in South America, their origin in South America and consequent dispersal from South America to Australia via Antarctica has been suggested 59. Additionally, this suggests one more case of strong faunistic affinities of the continent with South America and Australia 4, 6, 16, 60. So far, Antarctica has been considered as a dispersal route, but not as a probable place of origin. The new fossil finds support the hypothesis 10 that Antarctica may have acted as a center of diversification for australobatrachians.
The Seymour Island frog reported herein is the first vertebrate indicative of freshwater habitats on the Eocene Antarctic Peninsula, following invertebrate and plant evidence 12, 17 ( Fig. 6 View Figure 6 ). It is interesting to note that nearly all fossil localities where Calyptocephalella occurs (excepting those, for which fossil plant data are not available) contain evidence of the presence of Nothofagus , including Seymour Island 16, 60, 61. The southern extant range of Calyptocephalella occurs sympatrically with the microbiotherian marsupial Dromiciops gliroides ( Fig. S4 View Figure 4 ), also known as “Monito del Monte” or “Colocolo Opossum”, a small mammal with an arboreal lifestyle and an endemic
distribution in the dense Valdivian Nothofagus forests of highland Argentina and Chile 62. The climate of this Nothofagus forest area with the sympatric occurrences of these two endemic animals shows humid and temperate conditions (for the numerical values, see Methods and Table 2, Fig. S4 View Figure 4 ). Dromiciops gliroides is the only extant species of the order Microbiotheria and is considered as the only South American representative of the superor- der Australidelphia which otherwise comprises Australian marsupials 63. From the same small shell-rich lens that produced the frog remains reported herein, the fossil microbiotherian Woodburnodon casei has been described 64. Hence, we hypothesize that the climatic conditions for the Antarctic Peninsula during the Bartonian (late middle Eocene) should be comparable with the climate found today in the concurrent range of the Calyptocephalella- and Dromiciops -inhabited Nothofagus forests of South America.
The fossil finds of a frog and marsupial from Seymour Island, and their fossil and Recent distributions, represent outstanding examples of the role of global climate change on shifting biogeographic ranges. Despite global cooling and the disappearance of the habitats of these groups over large areas from Antarctica to Patagonia, they maintained their relictual occurrence in the Nothofagus forests of the central Chilean Andes. Thus, the Valdivian Nothofagus forest is a unique environment offering habitats not only for Eocene Antarctic refugees but also provides a modern analogue of the Antarctic climate just prior to the glaciation of the southern continent.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.