
Praxillinicola kroyeri M’Intosh, 1885
|
publication ID |
https://doi.org/10.11646/zootaxa.4174.1.22 |
|
publication LSID |
lsid:zoobank.org:pub:C7196500-B74B-423D-9FE1-3EB079B7F106 |
|
DOI |
https://doi.org/10.5281/zenodo.6071948 |
|
persistent identifier |
https://treatment.plazi.org/id/03AFBB1A-FFAC-FF97-0D8F-FB92FD7EFC4F |
|
treatment provided by |
Plazi |
|
scientific name |
Praxillinicola kroyeri M’Intosh, 1885 |
| status |
|
Praxillinicola kroyeri M’Intosh, 1885
( Figs. 7–8 View FIGURE 7 View FIGURE 8 )
Praxillinicola kröyeri M’Intosh, 1885 —incorrect original spelling (ICZN Art. 32.5.2.1).
Original description. M’Intosh (1885): 409, Plate XXXIX A, fig. 10.
Host. Praxillella abyssorum (M’Intosh, 1885) [as Praxilla abyssorum M’Intosh, 1885 ] (family Maldanidae ).
Type locality. Attached to incomplete specimen of P. abyssorum trawled at H.M.S. Challenger station 157 (Antarctic Ocean); 53º55’ S, 108º35’ E; depth 1,950 fathoms ( 3,566 m); diatom ooze. GoogleMaps
Material examined. Holotype ♀ (NHMUK reg. no. 85.12.1.305A); attached to the fifteenth segment of the holotype of P. abyssorum (NHMUK reg. no. 85.12.1.305) by “… two small processes on the under surface of the snout …” (M’Intosh 1885: 409); collected 03 March 1874.
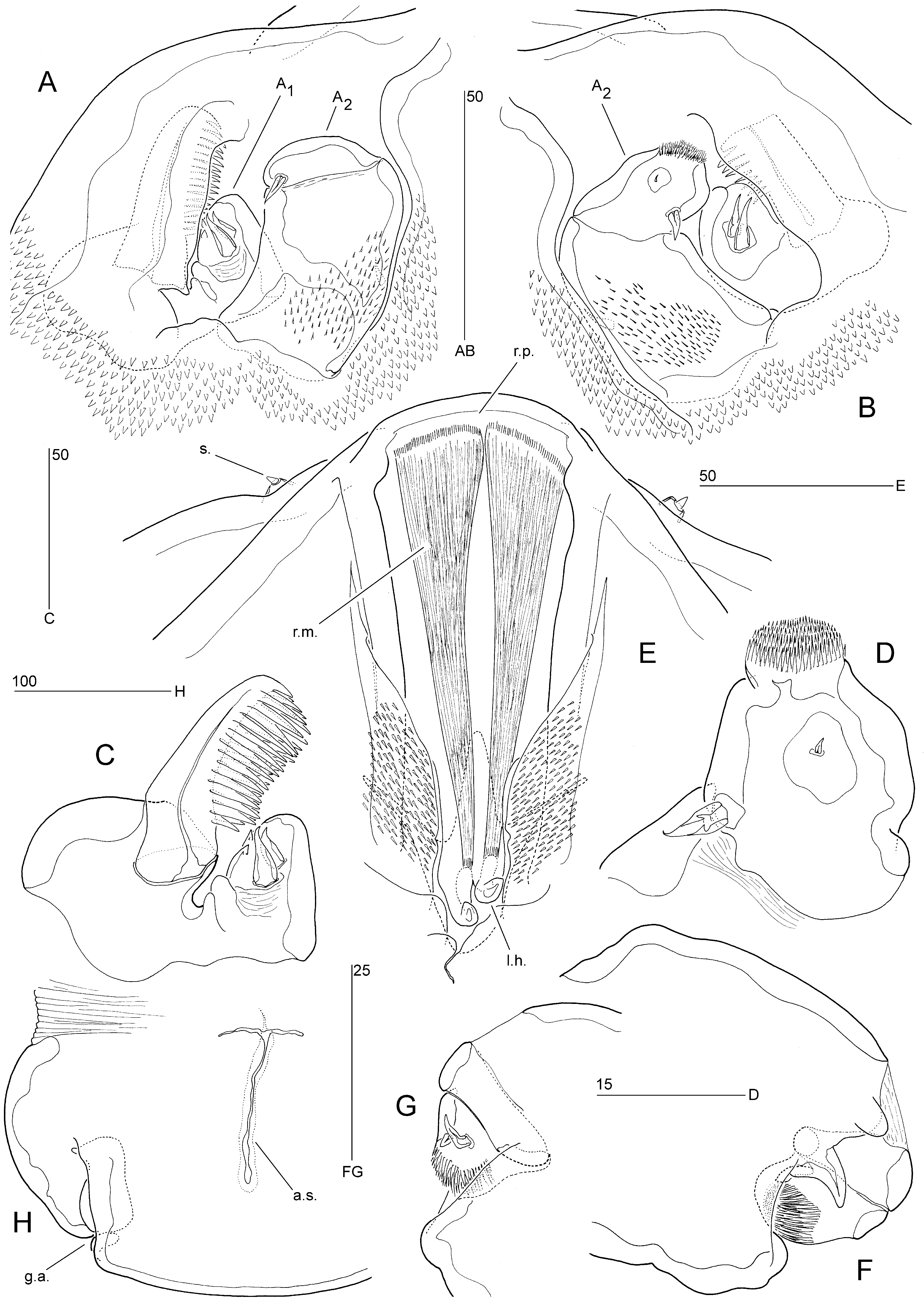
Redescription of female. Body elongate and dorsoventrally flattened ( Fig. 7 View FIGURE 7 A, B); consisting of cephalosome and indistinctly 5-segmented postcephalosomic trunk; total body length 2,190 µm, measured from anterior margin of rostral projection to posterior margin of abdomen (about 2.5 mm according to M’Intosh (1885)); maximum width 677 µm, measured halfway down the second postcephalosomic somite.
Cephalosome bell-shaped, separated from trunk by transverse membranous zone dorsally and ventrally ( Fig. 7 View FIGURE 7 A, B); dorsal surface with transverse furrow, median pore and four pairs of spiniform sensilla ( Fig. 8 View FIGURE 8 E). Surface of cephalosome with numerous denticles and spinules as indicated in Figs. 7 View FIGURE 7 A, B and 8A, B, E. Rostral projection arising from ventral surface, rounded, fused at base ( Figs. 7 View FIGURE 7 B; 8E).
Postcephalosomic trunk probably consisting of four thoracic prosomites and undivided urosome ( Fig. 7 View FIGURE 7 A, B); original somitic boundaries indicated by bilateral constrictions and transverse membranous zones dorsally and laterally; membranous zone between prosome and urosome continuous ventrally but interrupted middorsally by anal slit ( Fig. 8 View FIGURE 8 H). Prosomites limbless, with numerous spiniform and few setiform sensilla on dorsal surface as indicated in Fig. 7 View FIGURE 7 A; forming continuous unadorned sole ventrally ( Fig. 7 View FIGURE 7 B); fourth prosomite possibly compound as indicated by sensillar pattern. Capacious body cavity mostly filled with several hundreds of oocytes ( Fig. 7 View FIGURE 7 B).
Urosome 1-segmented, short, representing about 10% of total body length; about 1.5 times as wide as long; with paired dorsolateral genital apertures ( Figs. 7 View FIGURE 7 A; 8H) near posterolateral corners, no armature discernible; ventral surface with median depression in proximal quarter, containing paired copulatory pores (with minute associated pores posteriorly) ( Fig. 7 View FIGURE 7 C), and patch of minute spinules in distal quarter ( Fig. 7 View FIGURE 7 B). Anal operculum absent; anal slit triradiate ( Fig. 8 View FIGURE 8 H), positioned middorsally in anterior half of urosome. Caudal rami absent.
Ventral surface of cephalosome with paired anterior sockets containing vestigial antennules and antennae ( Figs. 7 View FIGURE 7 B; 8A, B). Antennule ( Fig. 8 View FIGURE 8 A–C) reduced, 1-segmented, consisting of large outer and smaller inner lobe; outer lobe with strong, medially directed, bipectinate claw; inner lobe with two short spines on posterior surface and two spiniform and two rounded extensions along outer margin.
Antenna ( Fig. 8 View FIGURE 8 A, B, D) 2-segmented, comprising coxobasis and 1-segmented endopod. Coxobasis without armature; posterior surface with numerous minute spinules. Endopod shorter than coxobasis; apex with spinular pad; posterior surface with one short and one minute spine.
Labrum ( Fig. 8 View FIGURE 8 E) narrow, produced into paired hook-like projections; three pairs of strong retractor muscles originating from frontal wall of cephalosome and inserting on bases of labral hooks (only one pair illustrated in Fig. 8 View FIGURE 8 E). Oral opening flanked by dense spinular patches. Paragnaths, mandibles, maxillules and maxillae absent.
Maxillipeds ( Fig. 8 View FIGURE 8 F, G) compact, located in distal third of cephalosome ( Fig. 7 View FIGURE 7 B); 3-segmented, comprising robust protopod (fused syncoxa and basis) and 2-segmented endopod. Protopod unarmed but with large spinous process on anterior surface, near articulation with endopod; forming concavity along palmar (medial) margin in which endopod can be withdrawn ( Fig. 8 View FIGURE 8 F). Enp-1 unarmed, partly fused with protopod along posterior surface ( Fig. 8 View FIGURE 8 G). Enp-2 narrower than enp-1; apex with spinular pad; posterior surface with two short spines.
Legs 1–6 absent.
Male. Unknown.
Remarks. M’Intosh’s (1885) description is very brief, illustrating only the whole animal in ventral aspect ( Fig. 1 View FIGURE 1 B). He interpreted the body as 9-segmented but it is not clear from his illustration which body regions his “segments” relate to. The abdomen was described as having two dilated somites anteriorly, followed by four narrower ones, the last one being the genital somite presenting a “...central genital aperture”. This study revealed that the postcephalosomic trunk probably consists of four thoracic prosomites and an undivided urosome. M’Intosh identified only one pair of appendages which he equated with the antennae (or antennules) with which the copepod attaches itself to the host. From their posterior position in his habitus drawing it appears that in reality he was referring to the maxillipeds. The central projection on the anterior ventral margin referred to as the rostrum by M’Intosh (1885) is here reinterpreted as the prominent labrum.
Praxillinicola kroyeri is unusual in the morphology and arrangement of its cephalosomic appendages. The paragnaths, mandibles, maxillules and maxillae are entirely absent while the strongly reduced antennules and antennae are contained within shared anterior sockets on the ventral surface of the cephalosome. Attachment to its maldanid host is secured by the maxillipeds, at least in females, and both the antennules and labral hooks probably function as auxiliary attachment organs. The distal apex of the maxilliped is blunt and modified into a distinct spinular pad as in other annelidicolous families such as the Anomoclausiidae , Clausiidae , Nereicolidae and Spiophanicolidae .
The antennule of Praxillinicola is the most outstanding character of the genus. The bilobate structure of the antennule with a strong, medially directed, bipectinate claw on the outer lobe has not been observed in any other genus of the Cyclopoida . The antennule of P. kroyeri shows a remarkable, but deceiving, similarity with the appendage tentatively identified as the antenna in Phyllodicola petiti ( Delamare-Deboutteville & Laubier, 1960a) . Laubier’s (1961) Fig. 3 View FIGURE 3 j (see also Delamare-Deboutteville & Laubier 1960a: Fig. C) shows a bilobate appendage with an outer branch consisting of two annular sclerites connected to each other by thinner areas of chitin (this condition was interpreted as 4-segmented by Boxshall & Halsey (2004)) and carrying two small claws at its tip, and an inner branch which resembles the bipectinate claw in P. kroyeri . However, Laubier’s illustration is an optical section through what appears to be a stacked series of semicircular lamellae originating from a common stalk (see his Fig. 3 View FIGURE 3 h). A similar structure was observed in the second phyllodicolid genus, Cyclorhiza Heegaard, 1942 ( Gotto & Leahy 1988) . Apart from their morphology being radically different, the structures also cannot be positional homologues since in Praxillinicola the claw originates from the outer lobe whereas in both phyllodicolid genera the lamellate structure represents the inner branch. Unlike P. kroyeri , adult females in the family Phyllodicolidae are mesoparasitic and utilize exclusively phyllodocid polychaetes as hosts ( Heegaard 1942; Delamare- Deboutteville & Laubier 1960a; Laubier 1961; Gotto & Leahy 1988; O’Reilly 2000).
Polychaete hosts are infested by a range of, mostly poecilostome, families ( Conradi et al. 2015). Approximately 170 species, representing 76 genera and 23 families are known to utilize polychaetes, either as ectoparasites, endoparasites or mesoparasites. None of the ectoparasitic members shows a comparable extreme reduction in body plan and segmentation as in Praxillinicola . Although the complete absence of legs 1–5 in the female is also displayed in some species of Anomopsyllus (Kim et al. 2013) and Vectoriella (Laubier & Carton 1973) , these nereicolids have an inflated trunk, a 2-segmented urosome and functional caudal rami, as well as a complete suite of cephalic appendages. Despite its highly transformed body P. kroyeri has retained the plesiomorphic condition of the female genital system, with the gonopores positioned dorsolaterally and the paired copulatory pores lying close together in a depression on the midventral surface. Among the poecilostome families this condition has been reported only in the Erebonasteridae ( Huys & Boxshall 1990; Humes & Huys 1992; Martínez Arbizu 1997). The presence of paired ventral copulatory pores was recently confirmed in the Serpulidicolidae (R. Huys, pers. obs.) and is probably related to the unusual ventral surface to ventral surface mating posture of the diminutive males in this family ( e.g. Southward 1964; Stock 1979). All serpulidicolids are external parasites of tubicolous, serpulid polychaetes. Although the hosts of most erebonasterids are currently unknown, at least one member is an associate of deep-sea vestimentiferan worms ( Humes 1973), a group that is now considered to belong to the polychaete family Siboglinidae ( Rouse 2001) . Some authorities ( Gotto 1979; Ho 1984) have considered the family Serpulidicolidae a member of the “Nereicoliform Group”, however, it is equally conceivable that it forms part of a different annelidicolous lineage including the Erebonasteridae and Praxellinicolidae. Maldanid polychaetes are typically, but not exclusively, utilized by members of the Clausiidae ( O’Reilly 1995; Kim et al. 2013), and occasionally by species of the Clausidiidae ( Bocquet et al. 1963) , but it is unlikely that P. kroyeri has a close affiliation to either of these families. Based on the unique characters discussed above it is here fixed as the type of a new family, Praxillinicolidae, which has independently entered into an ectoparasitic association with maldanid hosts.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |