
Bradophila pygmaea Levinsen, 1878
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4174.1.22 |
|
publication LSID |
lsid:zoobank.org:pub:C7196500-B74B-423D-9FE1-3EB079B7F106 |
|
DOI |
https://doi.org/10.5281/zenodo.6071957 |
|
persistent identifier |
https://treatment.plazi.org/id/03AFBB1A-FFB6-FF8E-0D8F-FF5CFE0BFD7C |
|
treatment provided by |
Plazi |
|
scientific name |
Bradophila pygmaea Levinsen, 1878 |
| status |
|
Bradophila pygmaea Levinsen, 1878
(Figs. 9–10, 11A)
Bradophila pygmaea Levinsen, 1878 —incorrect original spelling.
Bradophila pigmaea Levinsen, 1878 — Marchenkov (1999a: 108, 109, 111, 112, Figs. 34, 35; 1999b: 7, 8, 22, Table 1; 2002: 514, 516, 517): incorrect subsequent spelling.
Bradophyla pigmaea Levinsen, 1878 — Marchenkov (1998: 15; 1999b: 13, Table 3; 2001: 97; 2002: 517): incorrect subsequent spelling.
Bradophyla pigmae Levinsen, 1878 — Marchenkov (2001: 90, 97): incorrect subsequent spelling.
Levinsen (1878) discovered four females, of which two were ovigerous, among museum collections of the flabelligerid polychaete Brada villosa . His text description in old Danish and accompanying woodcut figures are remarkably detailed by contemporary standards and reveal details that were even overlooked in modern descriptions (e.g. Marchenkov 1999b, 2002). Contrary to Marchenkov’s (2002: 514) claim that the hosts were collected off the coast of Greenland by Steenstrup and Lütken no locality data were given (although note that some of the parasitic copepods described in Levinsen’s (1878) work did indeed originate from Greenlandic waters, e.g. Selioides bolbroei , Melinnacheres terebellidis ). According to Levinsen the egg sacs of ovigerous specimens extended into the “mouth cavity” of the host while the body was surrounded by, and probably embedded into, the anterior part of the annelid’s alimentary tract. Levinsen (1878) doubted whether he had dissected the entire parasite out of the host and his illustrations appear to confirm that he had only obtained the ectosoma. Hansen (1892, 1897) had previously suspected that some tubes must run into the body of the host, otherwise it would be difficult to see how the parasites obtain their food. The second report on B. pygmaea is that by Marchenkov (1997) who recorded a single ovigerous female from Brada villosa in the Chupa Inlet of the Kandalaksha Gulf, White Sea; no morphological observations were provided. In a conference abstract Marchenkov (1998) subsequently announced the discovery of additional material (four females and three males) from the same locality and host. He confirmed that the adult female body consisted of two portions, ectosoma and endosoma, connected by a well-developed stalk. Applying his newly defined criteria on host-parasite interrelations, Marchenkov (2001) provisionally considered Bradophila a potential mesoparasite pending the arrival of anatomical data.
Original description. Levinsen (1878): 371–374, Figs. xyl. (woodcuts) C–E.
Host. Brada villosa (( Rathke, 1843) (family Flabelligeridae ).
Type locality. Unknown. Levinsen’s material was found on polychaete specimens in museum collections, probably in the care of J.J.S. Steenstrup and C.F. Lütken.
Material examined. Damaged ovigerous ♀ with dwarf ♂ attached to ectosoma; obtained from B. villosa material collected in the Lofoten Islands and Norway and stored in The Natural History Museum ( NHMUK reg. no. 1900.5.1.364–366).
Notes on female. The following is based on Levinsen’s (1878) original description in Danish, Marchenkov’s (2002) Russian text and personal observations. Ectosoma almost spherical, slightly flattened in dorsoventral direction (on side adjacent to host); colourless or milky white in living specimens; measuring 500–700 µm in diameter ( Marchenkov 2002) (330–500 µm according to Levinsen); lacking all trace of segmentation or appendages. Paired genital apertures carried on highly sclerotized genital swellings at posterior end of ectosoma; located ventrolaterally and closed off by strongly chitinized, unarmed opercula. Single copulatory pore located between genital apertures; leading to seminal receptacle(s) via short copulatory duct. Stalk broad, campanulate (bell-shaped) in lateral aspect and passing through surface of host; anterior rim strongly chitinized, collar-shaped and folded backwards. Endosoma filiform, lying in body cavity along digestive tract of host; about 400–700 µm wide; as long as two-thirds body length of host; covered by numerous small rounded tubercles. Paired egg sacs multiseriate, elongate and disproportionately large, measuring 0.9–1.2 mm in length and 0.4–0.7 mm in diameter ( Marchenkov 2002) (1.50–1.66 mm long and ca. 0.75 mm wide according to Levinsen). Ectosoma with one or two males attached.
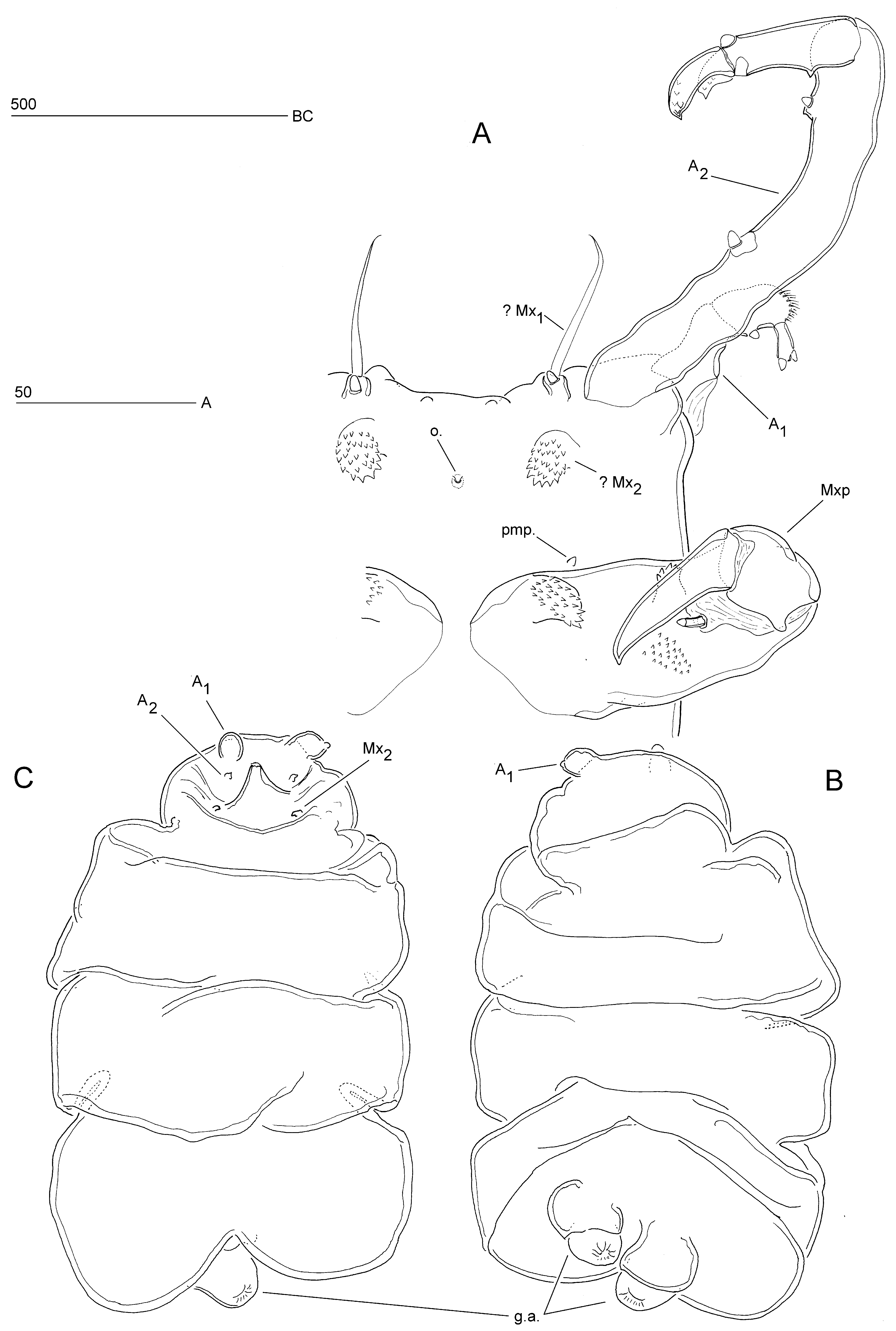
Redescription of male. Body (Figs. 9A; 10A) comprising broad, anterior cephalosome and slender elongate postcephalosomic trunk. Body length 530 µm, measured along dorsal curvature from anterior margin of cephalosome to posterior margin of caudal processes. Greatest width (93 µm) measured at cephalosome. Cephalosome (Fig. 9A) subquadrate in dorsal aspect; dorsal surface with pair of spiniform sensilla in anterior quarter; without surface ornamentation; anterior margin with paired lobate outgrowths dorsally (Fig. 9A) and smaller paired protuberances ventrally ( Fig. 11 View FIGURE 11 ). Trunk typically curved ventrally ( Fig. 10 View FIGURE 10 A); gradually tapering posteriorly in dorsal (Fig. 9A) but not in lateral aspect ( Fig. 10 View FIGURE 10 A); indistinctly 3-segmented; original segmentation marked by transverse membranous zones dorsally and laterally, and by vestiges of swimming legs 1–3 (arrowed in Fig. 10 View FIGURE 10 A). Caudal end with paired, medially directed, non-articulating, lobate processes (probably homologous to caudal rami); each process with spinules along outer margin and on posterior surface, and with dentiform projections on offset apical portion (Fig. 9D, E). Paired genital apertures located ventrally, just anterior to caudal processes (Fig. 9E); spermatophores paired.
Antennules short, located at anterolateral corners of cephalosome (Fig. 9A); short, with large, membranous insert at base dorsally ( Fig. 10 View FIGURE 10 A); 3-segmented ( Fig. 10 View FIGURE 10 C). Proximal segment largest, widest at base; distal anterior corner with blunt, posteriorly directed, styliform element. Middle segment subrectangular, positioned at right angle to segment 1; dorsal surface with two spinular patches and one modified conical element; posterior margin with modified rounded element. Distal segment smallest, slightly widening distally; directed at almost right angle to segment 2; carrying two modified elements apically.
Antenna ( Fig. 10 View FIGURE 10 B) uniramous, anteriorly directed (Figs. 9A; 10A); coxobasis and proximal endopod segment fused forming very long coxo-allobasis; free endopod indistinctly 2-segmented. Coxo-allobasis 3.9 times as long as endopod; with two modified blunt setae. Endopod with incomplete surface suture on ventral surface, indicating original segmentation; with one dorsal and one ventral modified blunt element; distal margin with two curved claws, each with two rows of dentiform projections.
Oral opening minute, probably not functional. Mandible absent. Homology of praeoral processes (Fig. 9B) difficult to assess. Anterior pair of processes (maxillules?) short and subcylindrical; located close to anterior margin of cephalosome, near bases of antennae; each with long naked seta and one modified blunt element (Figs. 9B; 11). Posterior pair of processes (maxillae?) lobate, backwardly recurved and rounded apically ( Fig. 10 View FIGURE 10 ); located either side of vestigial mouth; apical half covered with dentiform projections (Figs. 9B; 11). Pair of minute postmaxillary processes located anterior to maxillipeds (Figs. 9B; 11). Maxilliped (Figs. 9C; 10A) well developed, subchelate; comprising robust proximal segment (protopod) and 1-segmented endopod; palmar margin of protopod with two dentate processes (one proximal, one distal) on anterior surface, and denticular patch and one styliform modified seta on posterior surface ( Fig. 11 View FIGURE 11 ); endopod with strong curved terminal claw.
Remarks. While dissecting females on glass slides, Levinsen (1878) observed two small (200–250 µm) crustaceans, which he considered as belonging to a possibly early developmental stage of B. pygmaea . The dorsal view of this stage and accompanying text description leave no doubt that he was dealing with an adult male. According to Levinsen the instar consisted of an anterior, limb-carrying part and a slender, rounded posterior part which was about three times longer. He correctly identified the first pair of limbs as the antennules and described them as 3-segmented, ending in two thorns. The second pair of limbs was described as being the longest, having a large basal part followed by more, clear segments and ending in a thinner part, which also looked like it was segmented. Although this description is more difficult to reconcile with the observations reported here it is conceivable that Levinsen referred to the antennae. The fact that they were figured as posteriorly directed instead of forwardly is probably an artefact due to mounting. The third pair of limbs, which were illustrated as lateral lobes and described as consisting of one piece, which was thick and robust and provided with fine serrations along the anterior margin, can unequivocally be identified as the maxillipeds. Levinsen also observed remnants of a (spent?) putative male, presumably attached near the copulatory pore between the genital apertures of an ovigerous female. His statement that the only obvious parts were a long catching limb (“gribelem”) and a short, robust hooked limb (“kroglem”) makes a clear reference to the male antenna and maxilliped, respectively.
A fragmentary illustrated description of the male was presented in an unpublished Ph.D. dissertation ( Marchenkov 1999a, 1999b) in which it was suggested that B. pygmaea belonged in a separate monotypic family. The text description was eventually published as evidence in support of the proposal of the Bradophilidae ( Marchenkov 2002) while the illustrations were reproduced in Boxshall & Halsey’s (2004) treatment of the family. Unfortunately Marchenkov’s description contains several deficiencies and misinterpretations. His statement that B. pygmaea possesses dwarf males is misleading because the male (530 µm; 660 µm when antennae are included) is only slightly smaller than the ectosoma of the female (500–700 µm). The male attaches itself to the female by holding the stalk with its prehensile antennae and maxillipeds and using the caudal processes to grasp the ectosoma at the opposite pole. This posture ensures that the male genital apertures maintain close contact with the female copulatory pore, facilitating successful spermatophore transfer. Marchenkov (1999a, 2002) described the antennule as 4-segmented with the distal two segments being very small and unarmed; however, the apical segment in his drawing is in reality one of the two modified setae of segment 3. The long proximal segment of the antenna was interpreted as the coxobasis, however, the presence of two armature elements on this segment (poecilostome cyclopoids only have one) indicates that it is a compound segment derived by fusion of the coxobasis and the proximal endopodal segment, forming a coxo-allobasis. According to Marchenkov the mandibles are strongly reduced, consisting of a protruding basis, bearing one short seta near the outer margin, and a gnathobase represented by a thin long whip. Our re-examination showed that these limbs are not associated with the mouth opening and most likely represent the putative maxillules, the real mandibles being absent ( Fig. 11 View FIGURE 11 ). The maxillule, erroneously described as an indistinctly 2-segmented, slightly flattened, dentate lobe, in reality represents a different appendage, here identified as the putative maxilla. Marchenkov (1999a, 2002) described and illustrated the maxilla as a reduced lobe bearing small denticles along the inner margin; according to Boxshall & Halsey (2004) it is apparently 2-segmented, comprising a larger syncoxa separated from a tiny claw-like basis by a region of folded integument. Neither description is correct and the appendage labelled as the maxilla in Marchenkov’s (1999a) Fig. e could not be confirmed in the male examined herein. Paired minute postmaxillary processes were observed in the same position (Figs. 9B; 11) but it is unlikely that these structures represent remnants of the maxillae. Marchenkov (2002) described the maxilliped as 4-segmented but we could find no evidence for the alleged presence of a syncoxa.

| NHMUK |
Natural History Museum, London |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Bradophila pygmaea Levinsen, 1878
| Huys, Rony 2016 |
pigmaea
| Marchenkov 1998: 15 |