
Aporcelaimellus communis, Álvarez-Ortega, Sergio, Abolafia, Joaquín, Liébanas, Gracia & Peña-Santiago, Reyes, 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.214028 |
|
DOI |
https://doi.org/10.5281/zenodo.5679594 |
|
persistent identifier |
https://treatment.plazi.org/id/03B03611-8A06-FF95-FF4A-F9D80A11E01D |
|
treatment provided by |
Plazi |
|
scientific name |
Aporcelaimellus communis |
| status |
sp. nov. |
Aporcelaimellus communis sp. n.
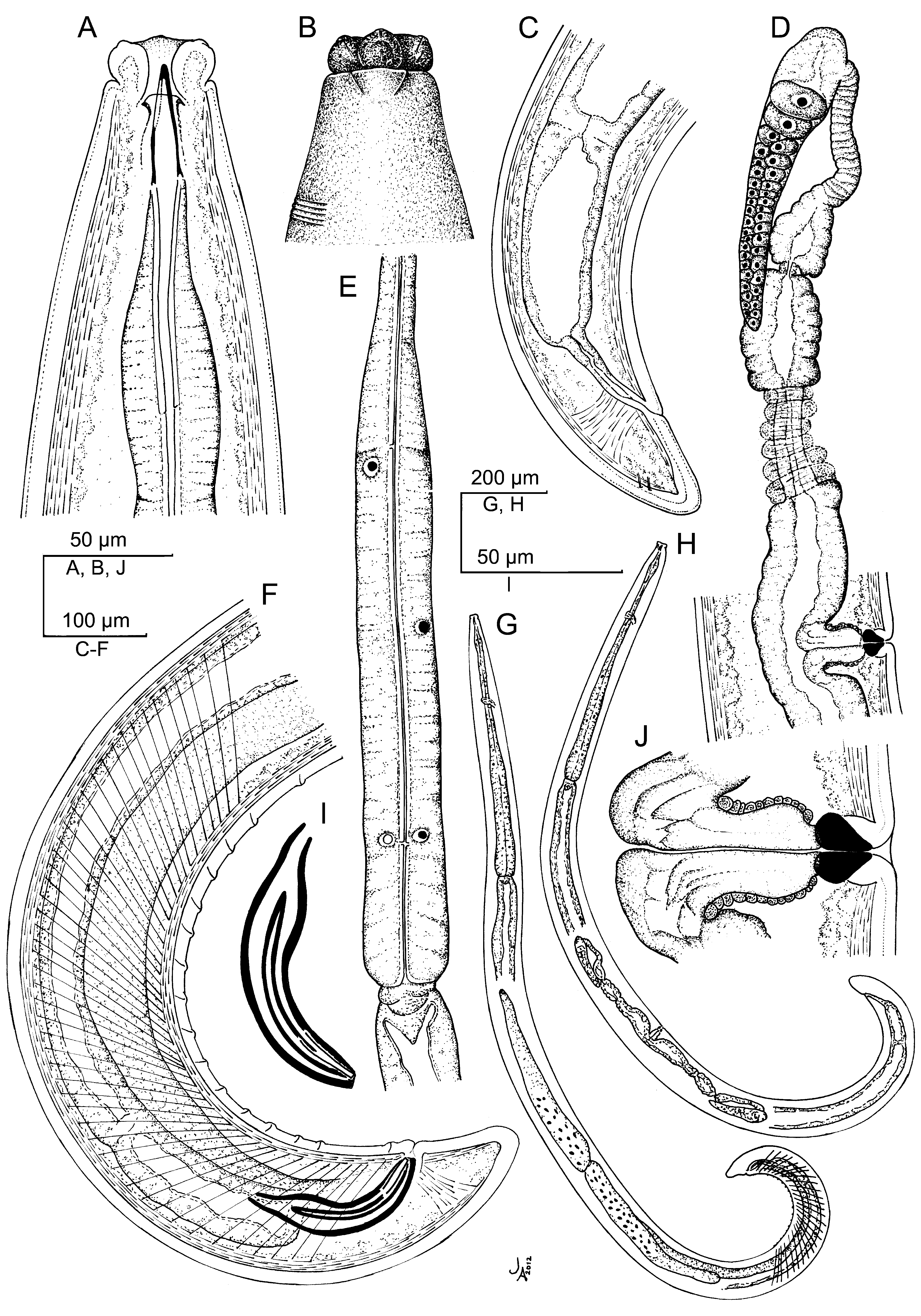
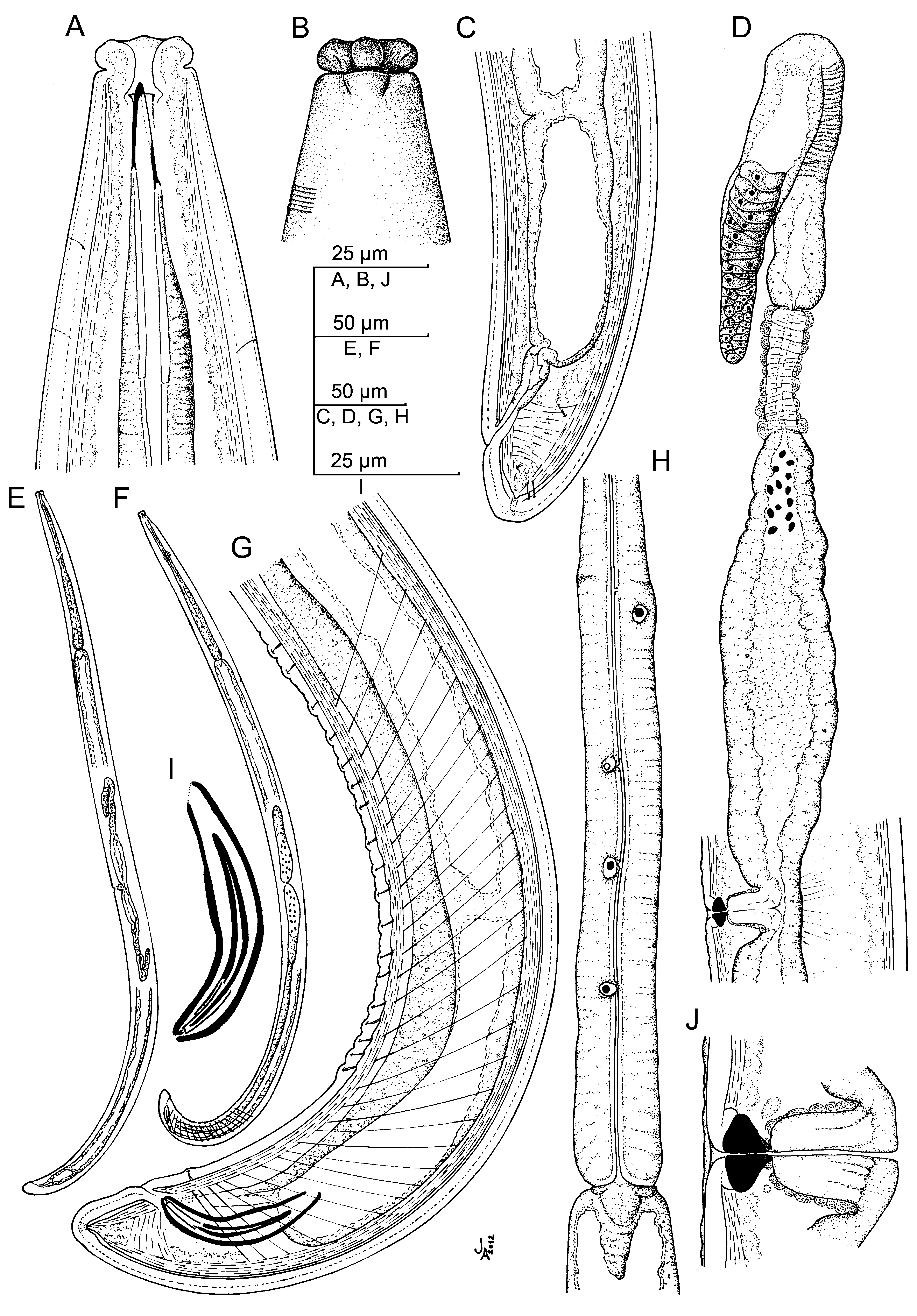
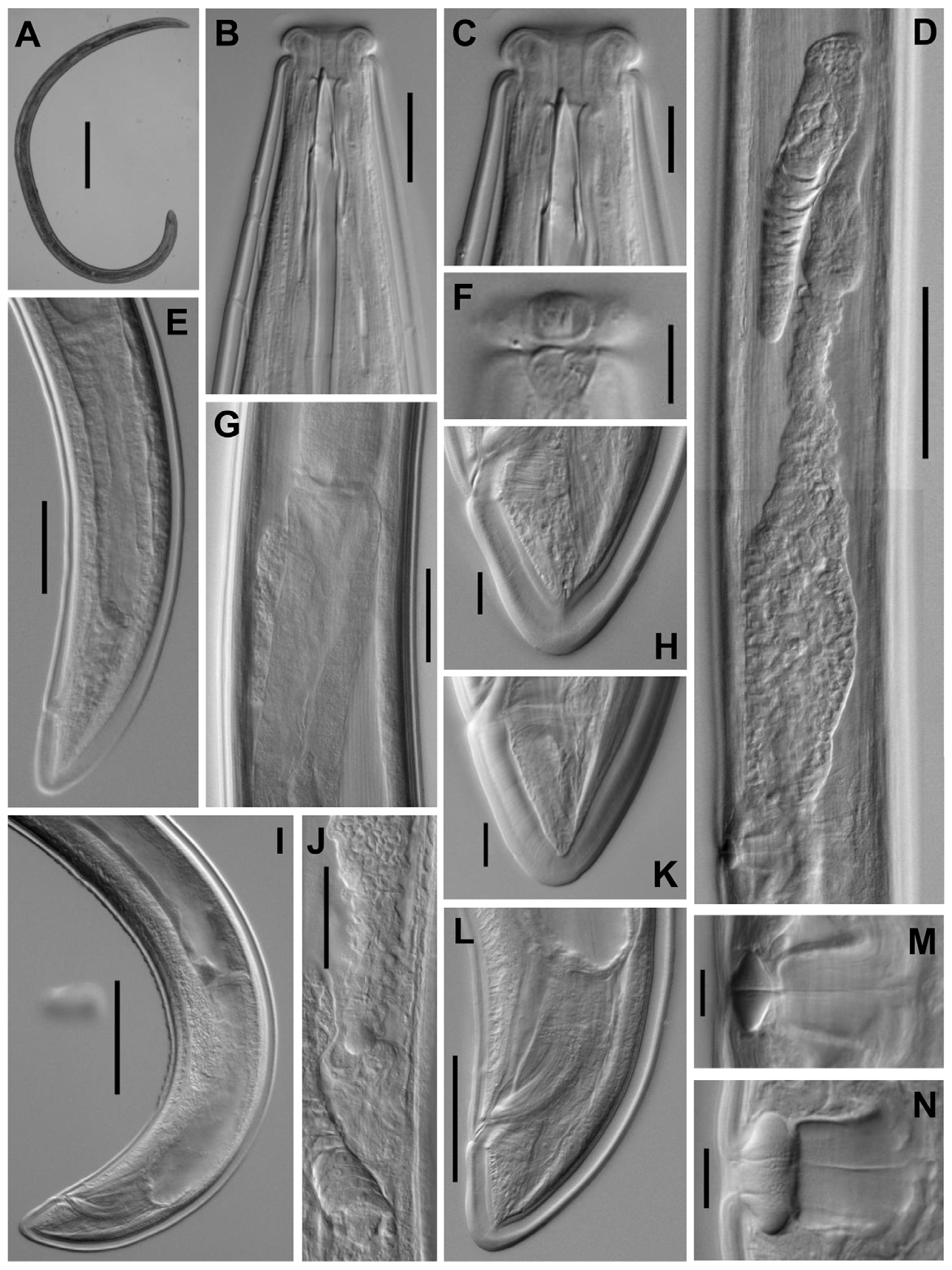
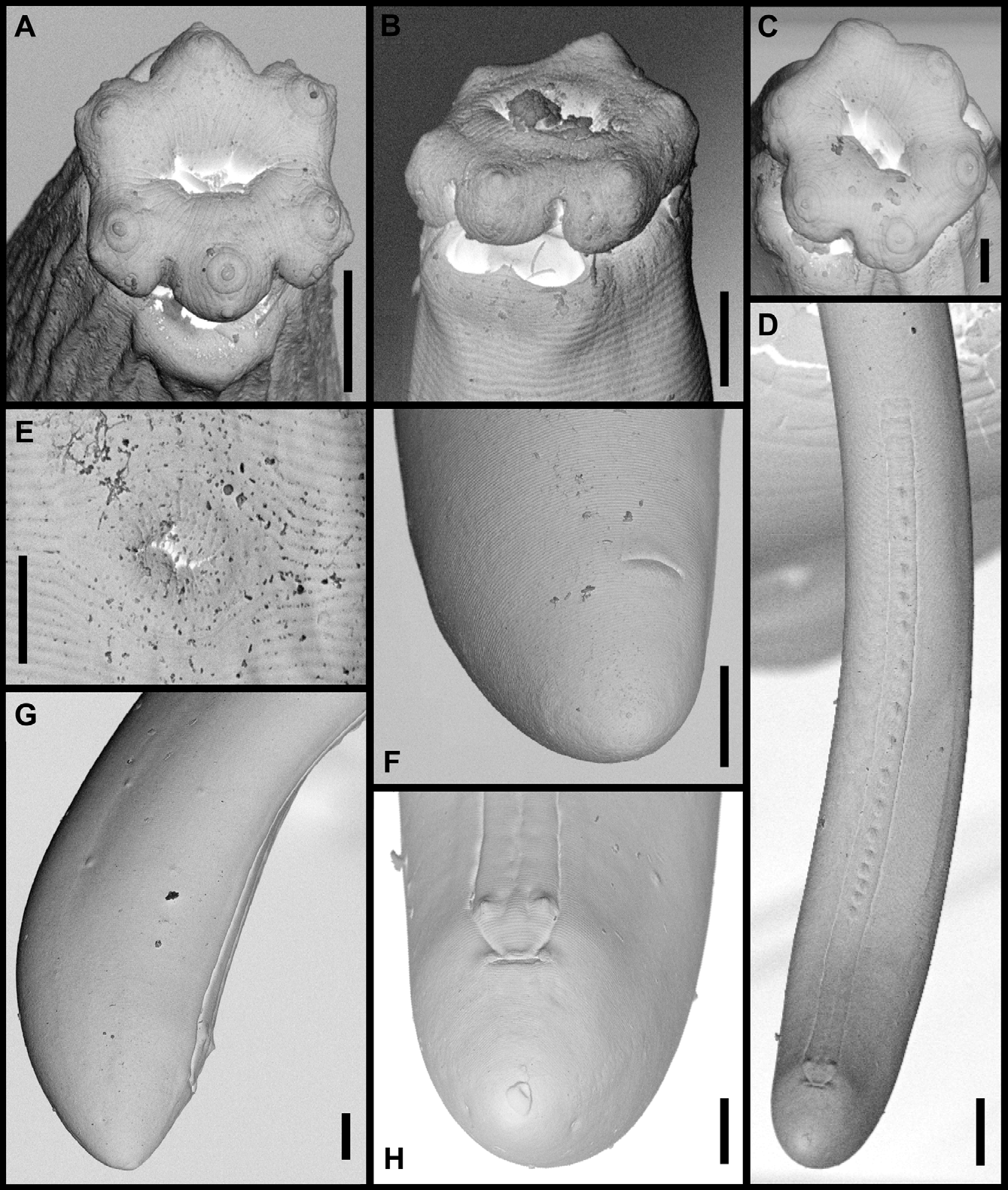
( Figs 5–7 View FIGURE 5 View FIGURE 6 View FIGURE 7 )
Material examined. Sixty-three females and 45 males from 19 localities of southeastern Iberian Peninsula; 24 females and 10 males in good state of preservation were measured.
Measurements. See Table 2.
Description. Adult: Moderately slender to slender nematodes of medium to large size, 2.56–4.22 mm long. Body cylindrical, tapering towards both extremities, but more so towards the anterior end. Habitus curved ventrad after fixation, especially in posterior body region, C-shaped, occasionally somewhat twisted. Cuticle three-layered, 2–4 μm thick at anterior region, 4–8 μm in mid-body and 6.5–12 μm on tail; outer layer thin and nearly smooth under LM but bearing very fine transverse striation under SEM, intermediate layer also thin, and inner layer thicker than the others and bearing radial striation. Lateral chord 7–17 μm wide or 7–16% of mid-body diameter. Two ventral and two dorsal body pores are usually present at level of odontostyle and odontophore, other body pores, more inconspicuous, are present at ventral side throughout the body. Lip region offset by deep constriction, 3.0–3.8 times as wide as high and one-fifth to two-sevenths (19–30%) of body diameter at neck base; lips (under SEM) mostly amalgamated excepting the lateral ones which are separated from the others by a deep incisure, hence the lip region shows a biradial symmetry; inner labial papillae surrounded by a ring-like annulus, while the outer labial and the cephalic papillae are not surrounded by any annulus; oral aperture a dorsoventral, hexagonal orifice, also contributing to the biradial symmetry of lip region. Amphid fovea funnel-shaped, distinctly protruding or bulging on body surface; its aperture 8–12 μm or three-sevenths to one-half (39–55%) of lip region diameter. Cheilostom nearly cylindrical, lacking any differentiation. Odontostyle typical of the genus, 4.2–4.9 times as long as wide, 0.9–1.1 times as long as lip region diameter, and 0.51–0.80% of body length; aperture 12–17 μm long or occupying two-thirds to three-fourths (64–74%) its length. Guiding ring plicate. Odontophore linear, rod-like, 1.9–2.3 times the odontostyle. Anterior region of pharynx enlarging very gradually; basal expansion 6.4–9.6 times as long as wide, 3.1–5.0 times as long as body diameter, and occupying 47–57% of total neck length; pharyngeal gland nuclei located as follows: DN = 52–58, S1N1 = 64–69, S1N2 = 75–78, S2N = 83–88. Nerve ring located at 185–229 μm from anterior end or 29–32% of total neck length. Cardia conical, 19– 42 x 18–29 μm; a well developed, ring-like structure is present surrounding its junction to pharyngeal base. A dorsal cellular mass about 100 µm long is very perceptible in many specimens at level of anterior end of intestine. Tail short and convex conoid, slightly more straight ventrally in males; inner cuticle layer almost reaching the tail tip, so that a hyaline portion is hardly distinguishable; caudal pores two pairs, one dorsal and other subdorsal.
Female: Genital system didelphic-amphidelphic, with large and equally developed branches, the anterior 340–693 μm or 11–19% of body length (521–800 μm or 16–23% of body length with uterine eggs inside of the uterus; 700 μm or 18% of body length with a egg inside of the oviduct and another inside the uterus), and the posterior 316–728 μm or 12–18% of body length (602–707 μm or 19–20% of body length with a egg inside of the oviduct; 641 μm or 16% of body length with a egg inside of the oviduct and another inside the uterus). Ovaries medium-sized, often reaching or shortly surpassing the sphincter level, the anterior 77–419 μm and the posterior 96–493 μm long; oocytes arranged first in two or more rows, then in a single row. Oviduct 114–300 μm long or 1.2–2.6 times the corresponding body diameter (257–340 μm long or 2.0–2.9 times the corresponding body diameter with one egg inside), consisting of slender part with prismatic cells and a moderately developed pars dilatata often with visible lumen. Oviduct and uterus are separated by a marked sphincter. Uterus long, bipartite, consisting of a slender distal region with narrow lumen, and a wider proximal part having thicker walls and distinct lumen; it is 190–450 μm long or 2.0–3.6 times the corresponding body diameter (313–382 μm long or 2.7–3.1 times the corresponding body diameter with a uterine egg inside); sperm cells are frequently found along the genital tract, but especially abundant at proximal region of uterus. Vagina extending inwards 31–53 μm or twosevenths to four-ninths (29–47%) of body diameter; pars proximalis 19– 37 x 16–28 μm, with somewhat sigmoid walls and surrounded by weak musculature; pars refringens with two triangular to drop-shaped, close (exceptionally separated) pieces measuring 7– 12 x 7–11 μm and with a combined width of 16–24 µm; pars distalis short, 1.5–4 μm. Vulva a short, oval, transverse slit; many females show cuticular irregularities near the vulva. Prerectum 1.7–3.9, rectum 0.9–1.4 times the anal body diameter.
Male: Genital system diorchic, with opposite testes. In addition to the adcloacal pair, situated at 16–28 μm from cloacal aperture, there is a series of 16–25 irregularly spaced (6–23 µm apart) ventromedian supplements, starting out the range of spicules, the posteriormost of which is situated at 63–134 μm from ad-cloacal pair. Two ventral, parallel incisures start a short distance in front of the anteriormost ventromedian supplement, end at cloacal aperture, and delimit a ventral band with the ventromedian supplements arranged on its mid-line. Spicules curved ventrad and slender, 5.4–6.9 times as long as wide, and 1.6–2.2 times as long as anal body diameter. Lateral guiding pieces 26–33 μm long, 7.0–8.3 times as long as wide. Prerectum 2.0–4.2, cloaca 0.9–1.7 times the anal body diameter.
Diagnosis. This species is characterized by its body 2.56–4.22 mm long, lip region offset by constriction and 19–25 μm broad, odontostyle 19–26 μm long with aperture occupying 64–74% its length, neck 595–750 μm long, pharyngeal expansion 321–427 μm long or 47–57% of total neck length, a dorsal cellular mass at level of anterior end of intestine, uterus bipartite and 190–450 μm long or 2.0–3.6 times the corresponding body diameter, pars refringens vaginae present, vagina a short oval transverse slit, V = 51–60, cuticular irregularities often present at vulva area, tail short and convex conoid (29–50 μm, c = 63–109, c’ = 0.5–1.0), spicules 89–118 μm long, and 16–25 irregularly spaced ventromedian supplements bearing hiatus.
Relationships. The new species is quite peculiar within the genus by the combination of a few characters: medium to large size, L = 2.56–4.22, uterus long and bipartite, and male with 16–25 irregularly spaced, ventromedian supplements.
It is very similar to A. pycnus ( Thorne, 1939) Baqri & Khera, 1975 , a rare species originally found in Hawaii although in plant material imported from Guatemala, and later only reported from Hungary by Andrássy (2002a) on the basis of female specimens. The new species is distinguishable from A. pycnus in a series of small but reliable and significant differences: comparatively shorter odontostyle (0.51–0.80 vs 0.87–1.02% of body length; data for A. pycnus calculated from Andrássy’s measurements) with much larger aperture (64–74 vs less than 60% of total length), more posterior position of S1N2 (distinctly closer to S2N than to DN [DN = 56, AS 1 = 27–29, AS 2 = 44–49 and PS = 65–70] vs distinctly closer to DN than to S2N [DN = 54–55, AS 1 = 17–18, AS 2 = 26–27 and PS = 66–70]), presence (vs absence, or not mentioned) of a dorsal cell mass at level of cardia, uterus with both sections of dissimilar length with slender portion much shorter than wider portion (vs both sections of similar length, see Andrássy’s original Fig. 12C; occupying 20–38% vs 42% of total uterus length), presence (vs absence or no mention) of cuticular irregularities near the vulva, prerectum without (vs with) a posterior blind sac, tail of both sexes lacking any dorsal concavity (vs a weak but distinct concavity always perceptible) and without a conspicuous hyaline portion (inner cuticle layer at tail nearly reaching the tail tip vs the hyaline portion of tail, typical of many Aporcelaimellus species, well distinguishable), and spicules more regularly and moderately curved ventrad (vs bearing a strong curvature, L-shaped, according with Thorne’s original description and Fig. 23b. Although these differences individually might be not especially important, together they conform to a pattern easily distinguishable from that known for A. pycnus (the type material for which is unavailable), and, being based on the study of many specimens, support the proposal of a separate status for the Iberian material herein examined.
In having medium to large size, comparatively wide lip region (19–25 µm) and long odontostyle (19–26 µm), the new species also resembles A. cocophilus ( Loos, 1949) Andrássy, 2001 , A. futaii , A. kazirangus Khan, Ahmad & Jairajpuri, 1995 , A. macropunctatus ( Heyns, 1967) Jiménez-Guirado, 1994 , A. micropunctatus , A. porcus and A. porosus . It differs from A. cocophilus in its shorter neck (vs 802–812 µm long), and male bearing hiatus (vs without hiatus, with five ventromedian supplements within the range of spicules); from A. futaii in its higher number (vs 7–10) and irregularly (vs regularly) spaced ventromedian supplements; from A. kazirangus in its larger general size (vs L = 1.9–2.3 mm, although male is said to be 3.4 mm long, certainly a mistake because according with original Fig. 3 View FIGURE 3 B both sexes are similar in size; neck 506–581 µm long), bipartite (vs simple) and longer (vs 163–168 μm) uterus, higher number (vs 11) and irregularly (vs regularly) spaced ventromedian supplements, and longer spicules (vs 72 µm); from A. macropunctatus in its larger odontostyle aperture (vs about one-half of total length), higher number (vs 10–14) and irregularly (vs regularly) spaced ventromedian supplements, and longer spicules (vs 68–80 µm); from A. micropunctatus in its bipartite (vs simple) uterus, higher number (vs 7–9) and irregularly (vs nearly regularly) spaced ventromedian supplements, and longer spicules (vs 79–87 µm long); from A. porcus in its female with vulva more posterior (V = 46) and male with higher number (vs 8) of ventromedian supplements; and from A. porosus in having less conspicuous (vs very distinct) body pores along the entire body, shorter odontostyle (vs 26–30 µm), bipartite (vs tripartite) uterus, and higher number (vs 8) of ventromedian supplements bering (vs lacking hiatus).
Type locality and habitat. Spain, province of Granada, near Bermejales reservoir, where it was collected associated with thyme ( Thymus sp.) and Ulex sp.
Other localities and habitats. Province of Almería: (i) Cabo de Gata-Níjar Natural Park, associated to Chamaerops humilis ; (ii) Sierra de los Filabres, associated with holm oak ( Quercus rotundifolia ); and (iii) Sierra Nevada-Alpujarras, associated with Retama sphaerocarpa , and thyme ( Thymus sp.). Province of Granada: (iv) Sierra Arana, associated with esparto (needle grass: Macrochloa tenacissima ) field; (v) Hoya de Baza, associated with Retama sphaerocarpa ; (vi) Vega de Granada, associated with holm oak ( Quercus rotundifolia ); (vii) Sierra Nevada Natural Park, associated with Retama sphaerocarpa , Ulex sp, juniper ( Juniperus sabina ), chestnut ( Castanea sativa ), and oak ( Quercus pyrenaica ); (viii) Sierra Nevada-Cerro Huenes, associated with Salix sp., pine forest ( Pinus sp.) and Ulex sp; and (ix) Sierra de Huétor Natural Park, associated with thyme ( Thymus sp.), holm oak ( Quercus rotundifolia ) and rockrose ( Cistus sp.). Province of Jaén: (x) Sierra de Cazorla, Segura y Las Villas Natural Park, associated with oak ( Quercus faginea ); (xi) Sierra de la Pandera, associated with Mediterranean brushwood, whose dominant species were Echinospartum boissieri, Salvia lavandulifolia, Lavandula latifolia and Erinacea anthyllis ; (xii) Sierra de Andújar Natural Park, associated with holm oak ( Quercus rotundifolia ), pine ( Pinus sp.) and cork oak ( Quercus suber ); (xiii) Despeñaperros Natural Park, associated with oak ( Quercus pyrenaica ) and fir forest; and (xiv) Sierra Mágina Natural Park, associated with maple trees ( Acer granatensis ), Ulex sp., holm oak ( Quercus rotundifolia ), pine trees ( Pinus halepensis ) and Mediterranean brushwood, whose dominant species were Erinacea anthyllis , Ptilotrichum spinosum , Vella spinosa , Ononis aragonensis and Astragalus granatensis . Province of Málaga: (xv) Sierra de las Nieves, (xvi) Valle del Genal, (xvii) Sierra del Pinar and (xviii) Sierra Bermeja, all of them associated with Abies pinsapo .
Type material. Female holotype and eleven female and three male paratypes, deposited in the nematode collection of the University of Jaén, Spain. Two female and one male paratypes deposited with USDA Nematode Collection, Beltsville, Maryland, USA.
Remarks. The abundant material examined of this species forms a rather homogeneous group from a morphological perspective. Nevertheless, some morphometric differences are noted between the specimens collected in the province of Jaén and those found in localities from the provinces of Almería and Granada: slightly broader lip region (23–25 µm, n= 8 vs 19–24, n=26, respectively), and longer odontostyle (23–27 vs 19–24 µm), odontophore (49–51 vs 39–45 µm) and spicules (99–118 µm, n= 4 vs 89–100, n=6). These differences are provisionally interpreted as intraspecific variability, but they might be the consequence of a process of geographical isolation of two groups of populations (possibly incipient subspecies).
Etymology. The specific epithet refers the high abundance of the new taxon.
| USDA |
United States Department of Agriculture |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.