
Hemibungarus calligaster (Wiegmann, 1834) , UTA
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2007.00350.x |
|
persistent identifier |
https://treatment.plazi.org/id/03B087FC-4C3E-FFD1-FCE9-70FB3B2D1DE1 |
|
treatment provided by |
Felipe |
|
scientific name |
Hemibungarus calligaster |
| status |
|
PHYLOGENETICALLY, HEMIBUNGARUS CALLIGASTER
IS NOT A CORALSNAKE
The taxonomy of Asian elapids, particularly the recognition of genera, has remained in flux for many decades, and has yet to be definitively inferred with strong well-supported phylogenetic evidence. Slowinski et al. (2001) were the first to use explicit phylogenetic methodology to estimate relationships among Asian and American coralsnakes including H. calligaster , although DNA sequence data for this species were unavailable and its relationships were assessed solely based on morphological data. Furthermore, their study included only a few non-coralsnake elapids (two for molecular and three for morphological phylogenetic analyses), and they constrained the monophyly of ‘coralsnakes’ (including Hemibungarus ), which was essentially necessary due to the limited inclusion of elapid outgroups in their study. However, this constraint had the effect of limiting possible alternative, non-traditional topological hypotheses to emerge from their analyses, including the phylogenetic clustering of H. calligaster with non-coralsnake elapines, rather than coralsnake genera, as we have found in this study.
In this study, although we have not included a majority of the (hydrophiine) genera of elapids, we have included a broad representation of essentially all known major elapid lineages (including all genera of elapines), as well as a diversity of colubroid outgroup taxa to avoid having to constrain elapid relationships (either explicitly or indirectly through limited outgroup inclusion). We found evidence that the coralsnakes exclusive of Hemibungarus ( Calliophis , Sinomicrurus , Micruroides , Micrurus and Leptomicrurus ) form a clade (ML-BSS = 68, PP = 96). Instead of being grouped with other Asian and American coralsnakes, ML and MCMC estimates place Hemibungarus with other elapine genera (ML-BSS <50, PP = 99). Hemibungarus forms a clade with Elapsoidea , Dendroaspis and Ophiophagus (ML-BSS <50, PP = 93), although support for relationships among members of this clade is weak ( Fig. 2 View Figure 2 ). A close relationship between Elapsoidea , Dendroaspis and Ophiophagus , and a distant relationship between these and other non-coralsnake elapines is generally similar to previous estimates ( Slowinski & Keogh, 2000). Our molecular phylogenetic results support the recognition of the monotypic Hemibungarus as a genus clearly distinct from Sinomicrurus or Calliophis (e.g. McDowell, 1987; Slowinski et al., 2001). Furthermore, our phylogenetic estimates suggests that H. calligaster is not phylogenetically a coralsnake, but rather shares an exclusive common ancestor with the Afro-Asian genera Ophiophagus , Dendroaspis and Elapsoidea .
MORPHOLOGICAL DISTINCTIVENESS OF HEMIBUNGARUS AND THE REINTERPRETATION OF HEMIPENIAL, COLORATION AND SCUTELLATION
CHARACTERS IN ELAPINE SYSTEMATICS
Molecular phylogenetic evidence presented here provides strong support for: (1) the phylogenetic distinctiveness of Hemibungarus calligaster , (2) a distant relationship between other coralsnake genera and Hemibungarus , and (3) evidence that the Elapinae comprises two main clades, one including exclusively coralsnake genera, and a second including noncoralsnake genera and Hemibungarus . In addition to molecular phylogenetic evidence, further evidence for these three conclusions is apparent when we reconsider the interpretation of hemipenial morphology, colour pattern and head scalation.
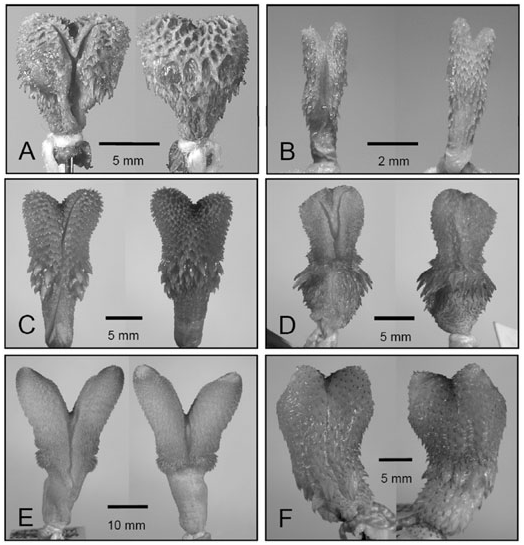
Examination of the hemipenes of exemplar Asian and African elapid species reveals substantial variation. The everted hemipenes of H. calligaster , Bungarus caeruleus , B. fasciatus , Dendroaspis polylepis , Naja naja and Ophiophagus hannah ( Figs 3 View Figure 3 , 4 View Figure 4 ) illustrate calyculation not present on the hemipenes of ‘true’ Asian coralsnakes (see Calliophis nigrescens , Fig. 3B View Figure 3 ) and also absent in American coralsnakes ( Slowinski, 1994; Campbell & Lamar, 2004; E. N. Smith, unpubl. data). This calyculation may be restricted to apical discs ( D. polylepis , Fig. 3F View Figure 3 ) or include only the tips of the organs ( N. naja , Fig. 3D View Figure 3 ), but appears to be always present in non-coralsnake elapine species. Contrastingly, the hemipenes of Asian coralsnakes of the genus Calliophis lack calyculation, and their only ornamentation are spines (see Fig. 3B View Figure 3 ; Slowinski et al., 2001). Slowinski et al. (2001), citing Leviton (1964), pointed out that McDowell (1986, 1987) was mistaken in reporting that H. calligaster possessed single (non-bifurcated) hemipenes, and here we confirm and illustrate the bilobed condition in H. calligaster . We have also found that the hemipenes of C. nigrescens are in fact bilobed, unlike C. gracilis , contra McDowell (1986, e.g. BMNH 98.4.2.27; fig. 5 in Slowinski et al., 2001 [AMNH 2870]). Additionally, we have observed that the hemipenes of both Calliophis species have no calyces (contrary to McDowell, 1986), and it appears that the hemipenes of all Asian coralsnakes ( Sinomicrurus and Calliophis spp. ) lack calyces. Collectively, the hemipenial character data suggest a close phylogenetic affinity of H. calligaster with non-coralsnake elapines, calyculate species.
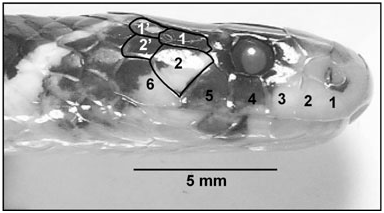
Slowinski et al. (2001) diagnosed H. calligaster , in part, as possessing a 1/1 temporal formula and a raised 6th supralabial; however, our observations lead us to an alternative interpretation. We interpret the raised sixth labial as a lower temporal of the first row, rendering the number of labials one less (five) and the temporal formula as 2/2 or 2/3 ( Fig. 5 View Figure 5 ). Leviton (1964) also regarded H. calligaster as having two primary temporals and six (rarely seven) supralabials. These character conditions are similar to those found in some other species of the Elapinae , particularly noncoralsnake elapine species that we estimate to be most closely related to H. calligaster using molecular data ( Fig. 2 View Figure 2 ). Many cobras and mambas possess an enlarged lower primary temporal, multiple secondary temporals and as low as six supralabials. Slowinski et al. (2001) also pointed out the uniqueness of the band arrangement in H. calligaster , consisting of black bands in dyads, as being a diagnostic character. The only other elapid that may possess this arrangement in the tail is Bungarus flaviceps baluensis from Borneo (see illustration in Vogel, 2006: 23), implying another potential link between Hemibungarus and noncoralsnake elapines.
TOWARD AN UNDERSTANDING OF RELATIONSHIPS
AMONG CORALSNAKES
As successive studies have added more resolution regarding relationships among Asian and New World coralsnakes, there are still a number of important (major) lineages that have yet to be confidently placed to frame inclusively the evolutionary history of coralsnakes. This study is, however, the first phylogenetic investigation to include one or more examples of all recognized genera of the Elapinae . Our ML and MCMC results provide strong support (ML-BSS = 88, PP = 99) for a clade consisting of temperate/tropical Asian ( Sinomicrurus ) and New World ( Micruroides , Micrurus and Leptomicrurus ) coralsnakes, and these estimates (unlike the MP estimates) are in strong agreement with morphological data. Specific morphological synampomorphic evidence for this grouping ( Sinomicrurus + American coralsnakes) has been identified: the loss of the postorbital bone ( McDowell, 1986), the presence of the basal pocket of the hemipenis ( Toriba, 1993), a uniformly spinose hemipenis ( Slowinski et al., 2001) and the bipartite origin of the m. adductor mandibulae externus superficialis muscle ( McDowell, 1986; Slowinski et al., 2001). Additionally, the lack of calyculation on the hemipenes might be considered a synapomorphic character, given that most non-coralsnake Elapinae and Hydrophiinae are calyculate. We also found moderate support (based on ML and MCMC) for the monophyly of American coralsnakes ( Fig. 2 View Figure 2 ; ML-BSS = 58, PP = 86), as have other studies based on molecular data ( Slowinski & Keogh, 2000; Slowinski et al., 2001).
Aside from the multiple forms of evidence linking Sinomicrurus with American coralsnakes, a clear understanding of the relationships of the other Asian coralsnakes ( Calliophis, sensu Slowinski et al., 2001 ) is lacking. Based on the musculature of the corner of the mouth, McDowell (1987) placed these remaining species into two groups: (1) Calliophis bibroni , C. gracilis and C. melanurus ; and (2) Maticora bivirgata , M. intestinalis , M. maculiceps and M. nigrescens . Slowinski et al. (2001) placed Maticora in the synonomy of Calliophis based on results of their morphological phylogeny estimate that arrived at a polytomy among members of these two genera. Our study, as well as the molecular data set of Slowinski et al. (2001), only included one species of Calliophis (C. [ Maticora ] bivirgata ) that we estimated to be the sister lineage of the remaining Asian and New World coralsnakes (ML-BSS = 66, PP = 96; Fig. 2 View Figure 2 ; as in Slowinski et al., 2001).
The morphological data set of Slowinski et al. (2001) does provide phylogenetic evidence that Calliophis (including Maticora ) is monophyletic, and the monophyly of Calliophis is supported by morphological synapomorphies: possession of a single elongate temporal scale, m. adductor mandibulae externus superficialis originating at the Harderian gland, and the posterodorsal extension of the Harderian gland ( Slowinski et al., 2001). These data suggest that all Calliophis species not included in this study form a clade with C. bivirgata , collectively implying that a monophyletic Calliophis forms the sister group to all other American and Asian coralsnakes. The hypothesis of a monophyletic coralsnake clade (excluding Hemibungarus ), and the validity of Calliophis (sensu Slowinski et al., 2001) , however, remain to be tested further with molecular data including expanded sampling of Asian coralsnake species.
Our sampling of New World coralsnakes included only a few representatives of this diverse group, although we have purposefully included a representative of each putative ‘major’ clade identified by Slowinski (1995). Our ML and MCMC estimates are congruent with nearly all previous studies based on molecular and morphological data in placing Micruroides as the sister lineage to all other New World coralsnakes (ML-BSS = 58, PP = 86; e.g. Roze & Bernal-Carlo, 1987; Slowinski, 1995; Slowinski et al., 2001; see also Gutberlet & Harvey, 2004). Our results also agree with Slowinski (1995; contrary to Roze & Bernal-Carlo, 1987) by providing well-supported evidence that Leptomicrurus renders Micrurus paraphyletic. Although Slowinski (1995) suggested the synonomy of Leptomicrurus with Micrurus , we have retained the use of the name Leptomicrurus (see also Campbell & Lamar, 2004) because this group of coralsnakes is distinctive (morphologically and genetically) and probably monophyletic ( Roze & Bernal-Carlo, 1987; Slowinski, 1995; Campbell & Lamar, 2004), and a thorough understanding of coralsnake relationships may eventually facilitate the dissection of the large genus Micrurus (~ 70 spp.) into multiple distinct genera (and the continued recognition of Leptomicrurus ). Recently, some of us (Smith, Parkinson, J. A. Campbell and Castoe) have initiated a large-scale investigation of relationships among Asian and New World coralsnakes. Our research continues to investigate the relationships among Asian Calliophis and Sinomicrurus , as well as the relationships among the New World coralsnakes using morphological and molecular phylogenetic data, and should eventually resolve many of the outstanding questions surrounding coralsnake evolution and systematics.
HIGHER- LEVEL RELATIONSHIPS AMONG ELAPIDS
A majority of authorities divide the Elapidae into two groups, the Hydrophiinae and Elapinae ( Hydrophiidae and Elapidae ), based largely on characters associated with the kinetic morphology of the skull ( McDowell, 1970), differentiating between the ‘palatine draggers’ and the ‘palatine erectors’. McDowell (1970) showed that the Afro-Asian cobras, Asian kraits, Asian and American coralsnakes, Laticauda and Parapistocalamus were ‘palatine erectors’ ( Elapinae ), and the remaining Australo-Paupan terrestrial elapids and sea snakes were ‘palatine draggers’ ( Hydrophiinae ). Subsequently, other studies have provided evidence for Laticauda being an early diverging lineage within the Hydrophiinae (palatine draggers, e.g. Cadle & Gorman, 1981; Schwaner et al., 1985; Slowinski et al., 1997; Keogh, Shine & Donnellan, 1998; Scanlon & Lee, 2004), implying that the palatine characteristics in Laticauda and probably Parapistocalamus ( Scanlon & Lee, 2004) may be either convergent or symplesiomorphic.
Based on analyses of our molecular data, some differences in phylogenetic signal for higher-level relationships among elapids were evident between the mtDNA and c-mos data set. Like the combined data analysis, the mtDNA data supported ( Hydrophiinae (coralsnake elapines, Afro-Asian non-coralsnake elapines)). The c-mos data suggested an alternative arrangement with the Hydrophiinae nested within the Elapinae , and a more basal divergence of coralsnakes: (coralsnake elapines ( Hydrophiinae , Afro- Asian elapines)). It is important to note, however, that we observed drastic variation in branch lengths based on c-mos across elapids, particularly among the hydrophiine genera, which may lead to inaccurate phylogenetic estimates when this locus is used as the sole data set for phylogenetic reconstruction.
The results of our combined data ML and MCMC analyses (and particularly the MCMC) support the existence of the Hydrophiinae and Elapinae as the two primary lineages of elapid snakes ( Fig. 2 View Figure 2 ). Support values for the monophyly of the Elapinae (ML-BSS <50, PP = 85) and the monophyly of the Hydrophiinae (ML-BSS <50, PP = 98) were low based on ML, but moderately to substantially higher based on the partitioned MCMC analyses of the combined data set ( Fig. 2 View Figure 2 ). We also found support for the Elapinae being composed of two main clades: the coralsnakes (excluding Hemibungarus ; ML-BSS = 66, PP = 96), and a second clade containing the Afro- Asian cobras, Asian kraits, and mambas (ML- BSS <50, PP = 99; Fig. 2 View Figure 2 ). This deep division among elapine snakes was also suggested by some of the results of Slowinski & Keogh (2000), and is compatible with the partially resolved phylogenetic topologies estimated from sequences of venom proteins ( Slowinski et al., 1997). This division of elapine taxa is also indicated by hemipenial characters (particularly the presence or absence of calyces as discussed above; see also Keogh, 1999).
Given the concordance of several lines of evidence (our phylogenetic estimates, previous phylogenetic estimates and hemipenial data), we suggest that the two main clades of elapines identified in ML and MCMC analyses be assigned taxonomic status. We suggest the name Hemibungarini for the clade encompassing the common ancestor of Hemibungarus ( Peters, 1862; designated as the type genus) and the Afro-Asian kraits, mambas and cobras ( Fig. 2 View Figure 2 ). These snakes are palatine draggers possessing calyculate hemipenes. We suggest the name Calliophini ( Calliophis, Gray, 1835 ; designated as the type genus) for the second of these elapine clades that contains the Asian and American coralsnakes. This second group consists of palatine draggers that possess no calyculation of the hemipenes as a synapomorphy. As such, both clades are assigned tribal status under the Elapinae . This new tribal-level taxonomy for members of the Elapinae should provide added convenience for future reference.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.