
Calacalles
|
publication ID |
https://doi.org/10.5281/zenodo.191696 |
|
DOI |
https://doi.org/10.5281/zenodo.6220766 |
|
persistent identifier |
https://treatment.plazi.org/id/03B0A514-281B-9D55-5189-9950FCF4D2B3 |
|
treatment provided by |
Plazi |
|
scientific name |
Calacalles |
| status |
|
Calacalles View in CoL relationships and subgenera
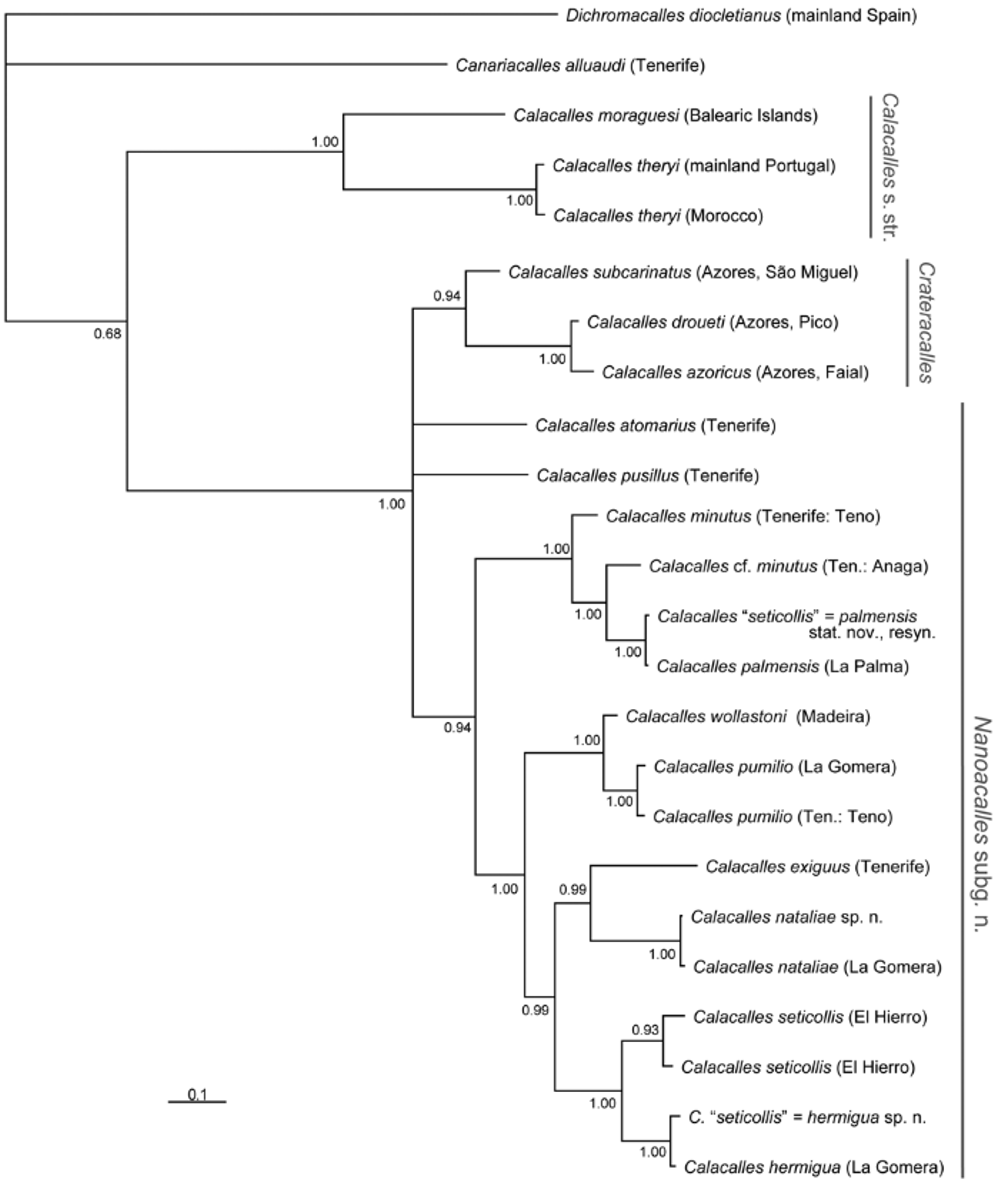
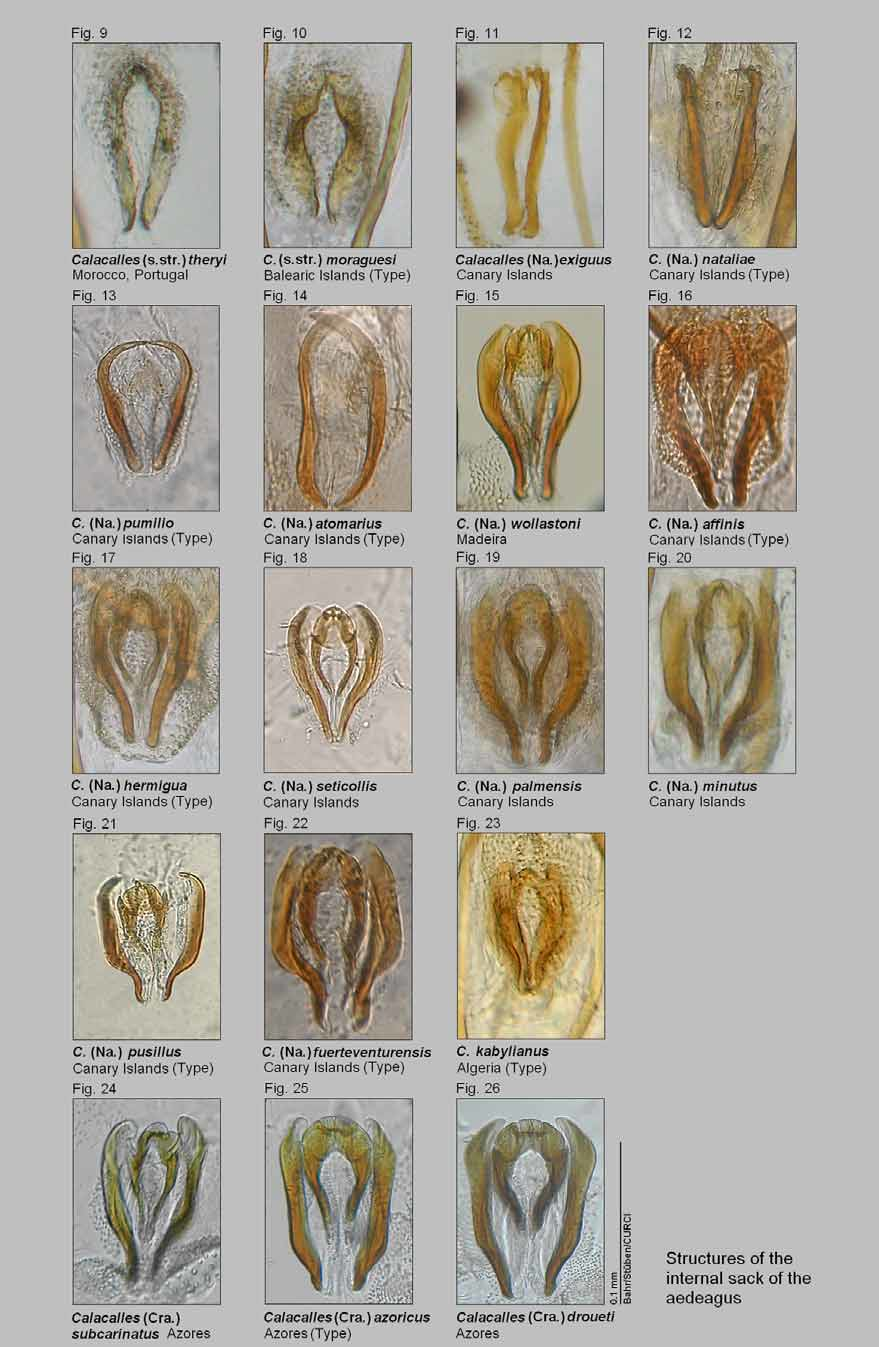
Among the usually tiny species of the genus Calacalles , exoskeletal differences are usually marginal. Equally—like in many other western Palaearctic Cryptorhynchinae—the female genital characters cannot be reliably used in species diagnosis (e.g. high intraspecific variability, different degree of sclerotisation of the spiculum ventrale and the spermatheca). For morphological analysis, only the male genitalia (the structure of the internal sac of the aedeagus, cf. Figs. 9–26 View FIGURES 9 – 26 ) offer the required stability (cf. Bahr 2000). In addition to the aedeagi, we here consider molecular genetic (mitochondrial) characters, which have not been used in Calacalles taxonomy before. Although we could not get hold of 3 of the so-far described Calacalles species for DNA sequencing, we see this paper (based on 15 out of 18 known Calacalles species) as a well-founded basis for a necessary revision including both morphological and molecular data.
Bodegas, N28 °33'38 " W16°0 9 '20",
500m, Sonchus spp., 2003, Stüben
Calacalles azoricus P.: Azores, Faial, Caldeira, N38°35'10" JJ0102, P-0125-azo EU286303 View Materials EU286468 View Materials
W28°42'04", 886m, Tolpis azorica, 2003 ,
Stüben
Calacalles droueti P.: A z o r e s, P i c o, P ic o d a U r z e, JJ0100, P-0123-dro EU286302 View Materials EU286467 View Materials
N 3 8 ° 2 7 ' 2 1 " W 2 8 ° 2 1 ' 0 4 ", 8 11 m,
Euphorbia stygiana, 2003 , Stüben
Calacalles exiguus E.: Tenerife, Anaga Mts., NE La Laguna, JJ0719, E-817-exi GQ332322 View Materials GQ332337 View Materials
Bosque de Las Mercedes, N28°31'49"
W 1 6 ° 1 7 ' 1 2 ", 9 0 5 m, L a u r u s
no vo c ana rie nsis, 200 8, Flor en &
Machado
continued next page Taxon Collecting data Vouchers Acc.# 16S CO1
Calacalles palmensis E.: La Palma, 11 km NE El Paso: Mt. JJ0098, C-0121-set EU286301 View Materials EU286466 View Materials
Tagoja, N28°43'18" W17°47'07", 1047m,
laurisilva, 2006, Stüben
Calacalles pumilio E.: La Gomera, S Hermigua , El Cedro: JJ0718, E-811-pum GQ332321 View Materials GQ332336 View Materials
Las Mimbreras, N28°07'27" W17°13'26",
901m, Myrica faya, 2008 , Astrin, Stüben,
Behne & Floren
Calacalles pumilio E.: Tenerife, Teno Mts., Monte del Agua, JJ0721, E-812-pum GQ332324 View Materials GQ332339 View Materials
N28°19'23" W16°49'15", 942m, Picconia
excelsa, Laurus novocanariensis, 2008 ,
Astrin, Stüben, Behne & Floren
Calacalles pusillus E.: Tenerife, Teno Mts., E Los Silos, El JJ0629, E-702-pus FJ716521 View Materials FJ716570 View Materials
Tanque Bajo, N28°21'39" W16°46'19",
534m, plant detritus, 2008, Astrin &
Stüben
Calacalles seticollis E.: El Hierro, 4 km SW La Frontera: El JJ0099, C-0122-set GQ332311 View Materials GQ332328 View Materials
Parque, N27°44'20" W18°01'30", 739m,
Sonchus hierrensis, 2006 , Stüben
Calacalles seticollis E.: El Hierro, 1 km SW Guarazoca: JJ0307, E-0265-set GQ332312 View Materials – C a m i n o d e l a Pe n a, N 2 7 ° 4 8 ' 0 6 "
W17°58'47", 746m, Carlina salicifolia,
2007, Stüben
Calacalles subcarinatus P.: Azores, São Miguel, Lagoa do Fogo, JJ0716, P-485-sub GQ332319 View Materials – a b o v e p o w e r p la n t, N 3 7 ° 4 6 ' 1 6 "
W25°29'07", 720m, Ilex perado, 2003 ,
Stüben
Calacalles theryi P.: N Caldas da Rainha, São Martinho do JJ0720, E-865-the GQ332323 View Materials GQ332338 View Materials
Porto, N39°30 '24" W0 9 °0 8'48 ", 3m,
Crithmum maritimum, 2009 , Astrin
Calacalles theryi M O R O C C O: 2 k m S E s s a o u i r a, JJ0724, E-914-the GQ332326 View Materials GQ332341 View Materials
N31°28'08" W09°45'27", 22m, Senecio
anteuphorbium at riverside, 2009, Stüben
Calacalles wollastoni P.:, Madeira, Santana, N 32° 48' 12" JJ0444, P-507-wol GQ332313 View Materials GQ332329 View Materials
W16°52'53", 457m, Castanea sativa,
2008, Stüben
OUTGROUP:
Canariacalles alluaudi E.: Tenerife, Teno Mts., Casa Blanca near JJ0622, E-695-all GQ332314 View Materials GQ332330 View Materials
Buenavista del Norte, N 28 °21 '36 "
W 1 6 ° 5 2 ' 1 0 ", 2 3 1 m, C e ro p e g i a
dichotoma, 2008, Astrin & Stüben
D ic h ro m a c a ll e s E.: Prov. Cádiz, 6,5 km S Algeciras, JJ0178, E-0204-dio EU286336 View Materials EU286500 View Materials
diocletianus P u n ta d e l C a r n e r o, N 3 6 ° 0 4 ' 3 5 "
W05°25'46", 34m, thistles, 2007, Astrin
& Stüben
At this point it should be mentioned that even though the exoskeletal characters of the Calacalles species look very similar at first view, it is the clues gained from molecular analysis that enable a more differentiated morphological investigation.
This does not mean that morphological analysis was 'dirtier' before applying DNA sequence analysis, but it is much easier, for example, to interpret variation as interspecific variation when we are able to base ourselves on molecular data. Such was the case in the 'discovery' of the new species Calacalles hermigua described below.
Synonymy of the subgenus Saetiacalles Bahr, 2000 : Both mitochondrial genes unambiguously suggest joining Calacalles moraguesi (funicle with 7 antennomeres) with Calacalles theryi (6 antennomeres), as proposed by Stüben (2005). During the last years, the number of antennomeres played a decreasing role in the taxonomy of Curculionoidea (e.g. Tychius , Ceutorhynchus ; Clark 1976; Caldara 1990; Colonnelli 1984). It can be argued that structures repeated serially in an organism, i.e. homonomous segments like antennomeres, are often controlled by a single gene. While phenotypically 'feigning' a macromutation, the variation in antennomeres is in fact probably nothing more than the product of repeated gene activity. Such segmental duplications or reductions are – phylogenetically – of lesser importance (cf. Wägele 2005 on homonomies or serial homologies).
Based on these theoretical considerations as well as on the above-mentioned molecular findings ( Fig. 1 View FIGURE 1 ), we reject a subgenus Saetiacalles Bahr, 2000 and abandon the use of the number of antennomeres as a taxonomic criterion. Instead, we propose a new subgeneric classification for the genus Calacalles as follows:
Calacalles s. str.: Figure 1 View FIGURE 1 shows the consensus tree of the phylogenetic analyses. The continental species Calacalles theryi and C. moraguesi are recovered as sister to all other Calacalles species, from which they are separated by very high genetic distances ( p - or uncorrected distance for e.g. C. moraguesi – C. atomarius : 10.9%). On the background of these molecular findings, we now also include Calacalles moraguesi with 7 antennomeres in the formerly monotypic subgenus Calacalles s. str. – misleadingly defined by Bahr (2000) as always possessing an antennal funiculus with 6 antennomeres (this change was already proposed by Stüben 2005: 123).
During typesetting of this article, we received another Calacalles sample from Libya that is morphologically very close to C. theryi , but molecularly distinct. It clearly belongs into the subgenus Calacalles s. str., but represents a distinct species from C. theryi and C. moraguesi . We suspect the sample to belong to the species C. kabylianus , of which only the damaged holotype from Algeria has been known so far. Therefore, in the Catalogue, we treat C. kabylianus as a Calacalles s. str. species. We will present the sequence of this sample in an upcoming publication.
Crateracalles Stüben, 2005: Sister to Calacalles s. str. stands a polytomy including, among others, an Azorean monophyletic lineage: the two large species of the subgenus Crateracalles plus the smaller C. subcarinatus . The two large Crateracalles species are nested within Calacalles , although morphologically, they are highly divergent. The two species are molecularly conspicuously close to each other (divergence of 1.1%) and it will have to be clarified whether they really represent two distinct species. The molecular and also new morphological evidence suggests the inclusion of C. subcarinatus into Crateracalles (see Taxonomy section).
Nanoacalles Astrin & Stüben subg. n. (see also Taxonomy section): Next to Crateracalles in the reconstruction stand the Canarian taxa: C. atomarius and C. pusillus by themselves, and the remaining species in a well-supported clade. This clade constitutes the new subgenus Nanoacalles, to which we also assign C. atomarius and C. pusillus due to their high morphological affinity to the other Madeiran-Canarian species. Hence Nanoacalles comprises the Madeiran-Canarian clade, but does not include the Azorean clade with C. droueti , C. azoricus , C. subcarinatus (all now Crateracalles) or the continental species of Calacalles s. str.: C. theryi , C. moraguesi , and C. kabylianus .
It can be noted that, according to our reconstruction, the Azores and the Canaries were colonised relatively early by Calacalles ( Nanoacalles) species, while Calacalles ( Nanoacalles) wollastoni from the
Madeiran archipelago evolved at a later time (this stands in contrast to the findings of Stüben & Astrin, in press, but the study included only six species of Calacalles ).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |