
Neochrysocharis beasleyi, Fisher, Nicole & Salle, John La, 2005
|
publication ID |
https://doi.org/ 10.5281/zenodo.169868 |
|
DOI |
https://doi.org/10.5281/zenodo.6266058 |
|
persistent identifier |
https://treatment.plazi.org/id/03B0EC45-FFEA-325E-FEC1-48D85BCF7DA3 |
|
treatment provided by |
Plazi |
|
scientific name |
Neochrysocharis beasleyi |
| status |
sp. nov. |
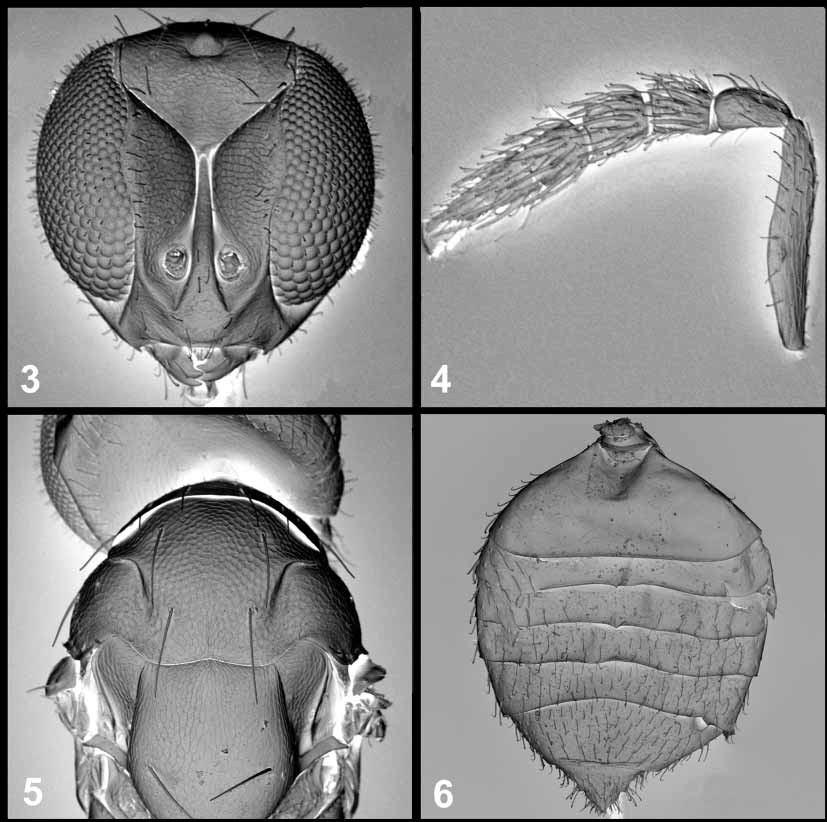
Neochrysocharis beasleyi , sp.n. ( Figs 1–6 View FIGURES 1 – 2 View FIGURES 3 – 6 )
Types. Holotype female: Indonesia, Bali, Bangli, Kintarnami, Buatian, coll. 7 Oct. 2001, I.W.Supartha, ex 10–17 Oct. 2001 on tomato, ANIC database no. 32 0 12622 (ANIC).
Paratypes. 64Ψ, 21ď paratypes as follows.
Indonesia: same data as holotype, (18Ψ 7ď: 13Ψ 4ď ANIC; 1Ψ 1ď BMNH; 1Ψ CNC; 1Ψ IZCAS; 1Ψ 1ď MZB; 1Ψ USNM; 1ď VAIC.) WJ – Bogor, Cisarua, coll. 21 Aug. 1997, A.Rauf, snowpea, ex. 28–30 Aug. 1997, tube no. 13 (3Ψ ANIC; 1ď USNM.) WJ – Bogor, Cisarua, coll. 8 Mar. 1998, A.Rauf, broccoli, ex. 10–18 Mar. 1998, tube no. 41 (1Ψ ANIC.) WJ – Bogor, Cisarua, coll. 21 Aug. 1997, A.Rauf, tomato, ex. 24–31 Aug. 1997, tube no. 42 (1Ψ ANIC.) WJ – Bogor, Cisarua, coll. 8 Mar. 1998, A.Rauf, broccoli, ex. 10– 18 Mar. 1998, tube no. 43 (1Ψ VAIC.)
Vietnam: Hung Yen Prov., My Van Dist, Vinh Khuc village, coll. 29 Sept. 1998, Tiep, bean, tube no. HY12 (2Ψ ANIC, 1Ψ BMNH, 1Ψ MZB, 1Ψ USNM); Hung Yen Prov., My Van Dist, Trung Trac village, coll. 10 Nov. 1988, Tiep, cucumber, tube no. HY18 (4Ψ ANIC, 1Ψ IZCAS); Hai Phong City, An Hai Dist, Tan Tien village, coll. 3 Nov. 1998, Hoa, tomato, tube no. HP34 (2Ψ ANIC, 1ď BMNH, 1ď CNC, 1Ψ IZCAS); Hung Yen Prov. My Van Dist, Tan Quang village, coll. 19 Sept. 1998, Tiep, cucumber, sample HY10 (1Ψ 2ď ANIC, 1Ψ CNC, 1ď IZCAS, 1ď MZB, 1Ψ USNM); Ha Nai City, Tu Liem Dist, Phu Dien village, coll. 13 Oct. 1998, Mai, on tomato, sample HN3 (1Ψ ANIC, 1Ψ 1ď BMNH, 1ď CNC); Ho Chi Minh City, Binh Chanh Dist, Vinh Loc A village, coll. 16 Aug. 1998, Ngoc, cucumber, sample HCM23 (1Ψ ANIC); sample HY2 (6Ψ 1ď ANIC, 1ď USNM); Hai Phong city, An Hai Dist., Tan Tien village, coll. 11 Nov. 1998, Hoa, cabbage, sample HP58 (1Ψ ANIC); HaNoi City, DongAnh Dist, Tien Duong village, coll. 3 Nov. 1998, Cuong, bean, tube no. HN12 (1Ψ ANIC); Hai Phong City, Thuy Nguyen Dist, Thuy Duong village, coll. 27 Nov. 1998, Hoi, chayote, tube no.HP50 (1Ψ ANIC); sample HCM4 (3Ψ ANIC); sample HPD5 (3Ψ ANIC); sample HY9 (1Ψ ANIC); Ho Chi Minh City, Binh Chanh Dist., Tan Tuc village, coll. 21 Aug. 1998, Ngoc, cucumber, sample HCM30 (1Ψ CNC); Hai Phong City, An Hai Dist, Hong Phong village, coll. 8 Sep. 1998, Hoa, long bean, sample HP23 (1ɗ IZCAS); Ho Chi Mnh City, Binh Chanh Dist, Tan Tuc village, coll. 3 Oct. 1998, Ngoc, jointed gourd, sample HCM14 (1Ψ MZB); Hai Phong City, An Hai Dist, Tan Tien village, coll. 3 Nov. 1998, Hoa, tomato, tube no. HP36 (2Ψ 1ɗ VA I C); Hung Yen Prov., My Van Dist, Trung Trac village, coll. 10 Nov. 1998, Tiep, cucumber, tube no. HP18 (1ɗ VA I C).
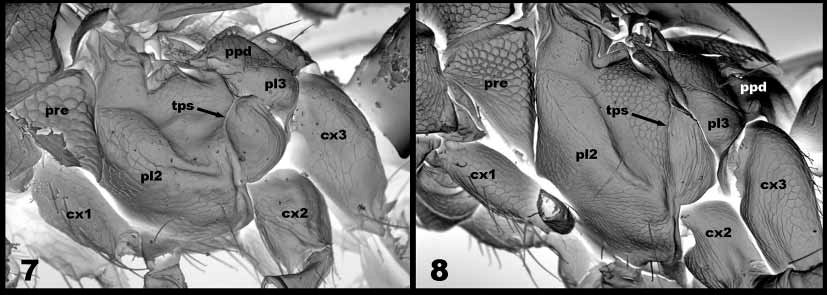
Diagnosis. Transepimeral sulcus (tps) curved posteriorly dorsally ( Fig. 7 View FIGURES 7 – 8 ). Female with all coxae dark metallic, in contrast to all femora and tibiae pale yellow to white; male similar, but hind femur with a distinct broad band of dark colour. Forewing completely hyaline, relatively broad: 2.0–2.15 times longer than wide. Mesosoma with weak reticulate sculpture, gaster relatively smooth and shiny. Coloration usually bluegreen metallic, occasionally with purplish reflections.
Description. Female ( Figs. 1–7 View FIGURES 1 – 2 View FIGURES 3 – 6 View FIGURES 7 – 8 ). Length of body 0.75–1.3 mm. Body metallic bluegreen, occasionally with purplish reflections. Legs: all coxae dark metallic, all femora and tibiae pale yellow to white.
Head ( Fig. 3 View FIGURES 3 – 6 ). Vertex, face and frons with weak to moderate reticulate sculpture. Frontal fork vshaped and distinctly separated from anterior ocellus.
Antenna ( Fig. 4 View FIGURES 3 – 6 ). Scape linear, 0.7–0.95 times longer than wide. Funicular segments about subequal in length and width. Club longer than funicle, segments 1 and 2 wider than long, segment 3 with terminal spine which is almost as long as the segment.
Mesosoma ( Figs. 5 View FIGURES 3 – 6 , 7 View FIGURES 7 – 8 ). Pronotum, mesoscutum and scutellum with weak reticulate sculpture dorsally. Pronotal collar without transverse carina. Midlobe of mesoscutum with 2 strong pairs of setae. Notaulus becoming broad and shallow posteriorly. Transepimeral sulcus (tps) curved posteriorly dorsally ( Fig. 7 View FIGURES 7 – 8 ). Propodeum smooth, shiny and without median carina; callus with 2 setae.
Forewing ( Fig. 2 View FIGURES 1 – 2 ) 2.0–2.15 times longer than wide; completely hyaline with large speculum. Marginal vein 4.0–5.25 times longer than stigmal vein; postmarginal vein 0.75– 1.2 times as long as stigmal vein. Cubital row of setae complete to basal vein; subcubital vein of setae indistinct and only present basally (distal to basal vein).
Gaster ( Fig. 6 View FIGURES 3 – 6 ) ovate. Gaster and thorax relatively smooth and shiny, with no distinct reticulate sculpture.
Male Length of body 0.6–1.15 mm. Male similar to female, but hind femur with a distinct broad band of dark colour.
Biology. No specific biological studies have been performed on this species. Like other Neochrysocharis , it is presumed to be an idiobiont endoparasitoid of host larvae.
Distribution. This species is only known from Indonesia and Vietnam, but is probably widespread through Southeast Asia.
Etymology: This species is named in honour of Edmund “Ted” Beasley, a good friend of ANIC, and a long time volunteer in the collection.
Discussion. Neochrysocharis species are endoparasitoids of a variety of small phytophagous insects, mainly leafminers on herbaceous plants, but other hosts are known. There has been some controversy as to the status of the generic name Neochrysocharis . Boucek (1988) considered Neochrysocharis as a synonym of Chrysonotomyia Ashmead , although Hansson (1990) treated the two genera as distinct and supplied a key to European species of both genera. Hansson (1994) subsequently recognized Chrysonotomyia as containing only a few New World species, with the bulk of the world species that had been treated in this genus properly belonging in Closterocerus Westwood. He has supplied revisions of species of North American Closterocerus ( Hansson, 1994) and Neochrysocharis ( Hansson, 1995) . These works offer more information on the taxonomy and biology of these insects.
Gumovsky (2001) synonymised Neochrysocharis , Asecodes Delucchi, Hispinocharis Boucek and Mangocharis Boucek with Closterocerus . Further study will be required to gain a complete understanding of relationships in these groups; until that time, we are considering Neochrysocharis and Closterocerus as separate genera. Neochrysocharis stat. rev. currently contains about 45 species, and is cosmopolitan in distribution ( Noyes, 2002; 2003).
There are no keys available for Southeast Asian species of Neochrysocharis . Using the key for Palaearctic species ( Hansson, 1990), N. beasleyi would key to couplet 13. At couplet 13, N. beasleyi no longer fits the key, and differs from remaining species as follows: female with all coxae dark metallic, all femora and tibiae pale yellow to white; male similar, but with the hind femur with a distinct broad band of dark colour. Additional characters for separating this species are given in the diagnosis.
Using the key to Nearctic species of Neochrysocharis ( Hansson, 1995) , N. beasleyi would key to couplet 4, based on having the tps curved dorsally and directed backwards from base of mid coxa. At couplet 4 it would differ from both remaining species (chalybea Hansson, texensis Hansson), by the coloration of the legs and strength of the sculpture on the mesosoma. Neochrysocharis beasleyi appears similar in habitus to the North American N. diastatae (Howard) . It differs from N. diastatae in having the dorsal portion of the tps curved posteriorly ( Fig. 7 View FIGURES 7 – 8 ), and in having all the femora and tibiae completely pale yellow to white; N. diastatae has the tps straight (similar to what is seen in N. formosa , Fig. 8 View FIGURES 7 – 8 ), and the legs generally with some dark coloration.
Keys for separating the three species of Neochrysocharis known to attack serpentine leafminers in Southeast Asia are given by Fisher et al, 2005. Briefly, N. beasleyi can be separated from N. formosa by having the forewing completely hyaline (forewing with an infuscate cloud near the stigma in N. formosa ); and from N. okazakii by having the forewing 2.0–2.15 times longer than wide (more than 2.5 times longer than wide in N. okazakii ). All three species have the coxae dark and the legs predominantly if not completely white to yellow. In this respect, Southeast Asian specimens of N. formosa differ from the typical European specimens of this species. This is discussed further in the section below on Southeast Asian Neochrysocharis formosa .
An additional character to separate these three Southeast Asian species is the shape of the transepimeral sulcus (tps). The tps is generally straight in Neochrysocharis , with its dorsal apex directed forward from the base of the mid coxa. This is the condition found in N. formosa ( Fig.8 View FIGURES 7 – 8 ). In N. beasleyi ( Fig. 7 View FIGURES 7 – 8 ) and N. okazakii the tps is curved dorsally, with its apex directed backwards from the base of the mid coxa.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |