
Cynelos anubisi, Morlo & Miller & Bastl & Abdelgawad & Hamdan & El-Barkooky & Nagel, 2019
|
publication ID |
https://doi.org/ 10.5252/geodiversitas2019v41a21 |
|
publication LSID |
urn:lsid:zoobank.org:pub:F2985B60-626C-4FF6-8C0F-C62C34531199 |
|
DOI |
https://doi.org/10.5281/zenodo.3703956 |
|
persistent identifier |
https://treatment.plazi.org/id/EED887B0-9F1C-4ADD-B4CF-7E8BBEB7514D |
|
taxon LSID |
lsid:zoobank.org:act:EED887B0-9F1C-4ADD-B4CF-7E8BBEB7514D |
|
treatment provided by |
Valdenar |
|
scientific name |
Cynelos anubisi |
| status |
sp. nov. |
Cynelos anubisi n. sp.
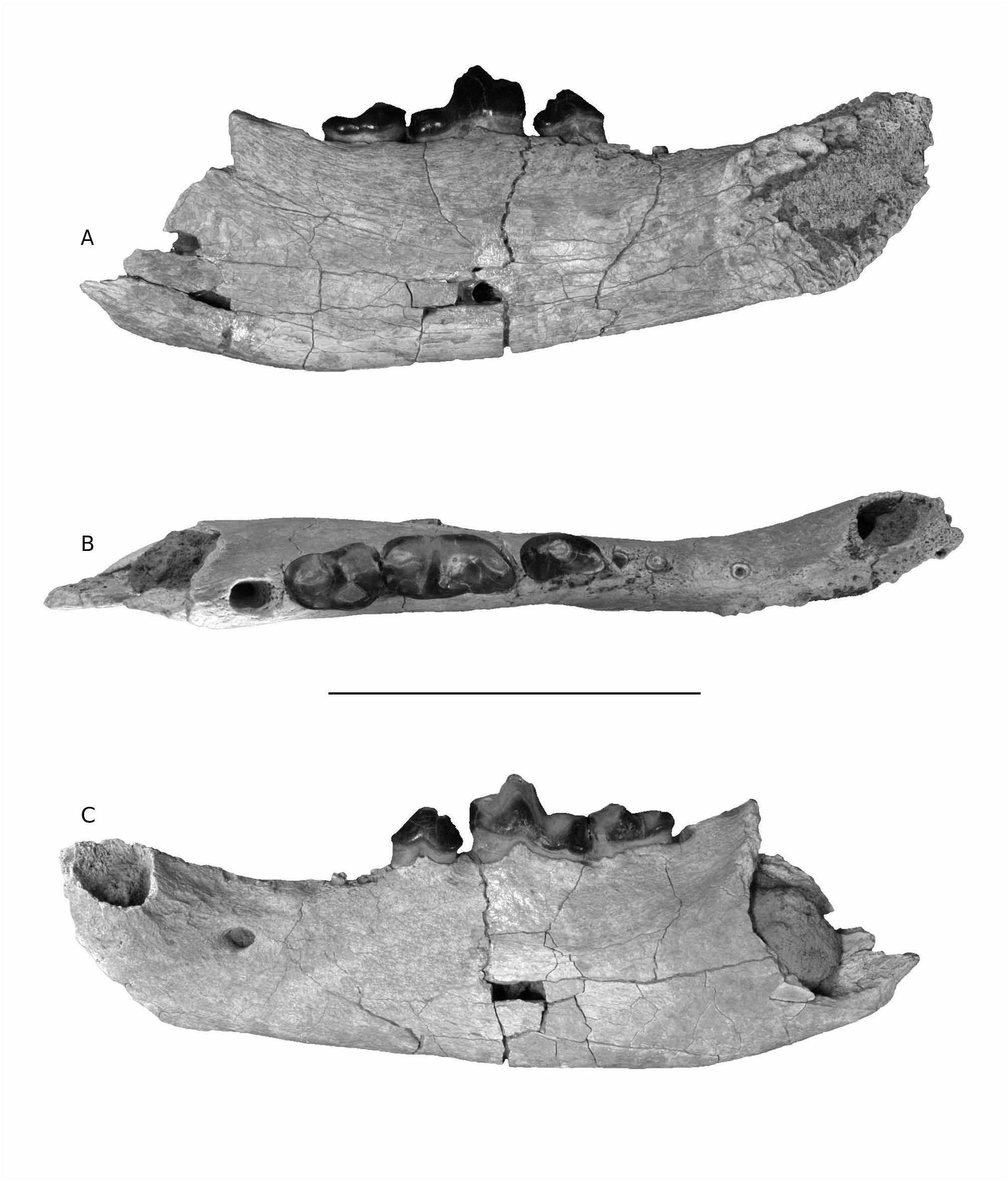
( Fig. 2 View FIG )
Cynelos sp. nov. – Morlo et al. 2007: fig. 4G, H.
cf. Ysengrinia ginsburgi – Morales et al. 2010: 48.
Afrocyon burolleti – Morales et al. 2010: fig. 3.
Amphicyon sp. – Morales et al. 2010: fig. 4.
Cynelos sp. – Werdelin & Peigné 2010: 604.
Afrocyon n. sp. – Morales et al. 2016: 143.
HOLOTYPE. — CUWM 55 View Materials , left mandible with alveolus of c, single root of p1, single root of p2, roots of p3, p4-m2, and alveolus of m3.
PARATYPE. — DPC 14532/2 L-7, isolated left m1 from Moghra, Egypt ( Morlo et al. 2007: fig. 4G, H).
REFERRED MATERIAL. — MNCN 79042 right mandible fragment with alveolus of p1, single-rooted p2, fragment of p3, and p4 from Gebel Zelten, Libya (= BIZ.2A. 15 in Morales et al. 2010: fig. 4). NHM M 82374, an isolated left m2 from Gebel Zelten, Libya ( Morales et al. 2010: fig. 3).
TYPE LOCALITY. — Wadi Moghra, Egypt.
DISTRIBUTION. — North Africa: Egypt and Libya.
AGE AND HORIZON. — Late Early Miocene, contemporaneous with European biozone MN4.
ETYMOLOGY. — “ anubisi ” after “Anubis”, the Greek name for the Ancient Egyptian jackal-headed god, who watched over the dead.
DIFFERENTIAL DIAGNOSIS. — Cynelos anubisi n. sp. differs from Amphicyon and Afrocyon in having a single-rooted p1 and p2, a long diastema between p2 and p3, and a short diastema between p3 and p4. The m1 metaconid of C. anubisi n. sp. is more reduced, the hypoconid is smaller, and the m1 and m2 talonids are narrower. The m2 protoconid and hy- poconid are low but massive. Cynelos anubisi n. sp. further differs from Amphicyon in having a narrower m2, as a result of an elongated talonid. All molars lack cingulids. Further differs from Afrocyon in having a single-rooted m3, and a coronoid process that rises at a shallower angle. Cynelos anubisi n. sp. is about the same size as, or is slightly smaller than C. macrodon, but C. anubisi n. sp. differs from C. euryodon and C. minor in being much larger in size, and differs from the Buluk Cynelos in being much smaller. Cynelos anubisi n. sp. differs from C. euryodon and C. lemanensis in having a single rather than a double rooted p2, and a much shorter p3-p4 diastema (see Peigné & Heizmann 2003; Morales et al. 2016); differs from C. macrodon and C. ginsburgi n. comb. in having m1 with a longer paraconid blade relative to overall tooth size; further differs from C. macrodon in having the m1 paraconid oriented slightly more anteriorly, having p4 smaller, and in possessing small p4 pre- and postprotoconulids; and further differs from C. ginsburgi n. comb. in having p4 and m2 longer relative to m1, m1 broader the m1 paraconid less angled but relatively longer, and the metaconid is located slightly more anteriorly. Cynelos anubisi n. sp. is larger than any Eurasian Cynelos species, including C. bohemicus ( Schlosser, 1899) which is known from Europe and Asia during biozones MN 3 to MN 5 ( Jiangzuo et al. 2018). Cynelos anubisi n. sp. also clearly differs from C. bohemicus in possessing a large diastema between p2 and p3 that is absent in this Eurasian species (see Fejfar & Heizmann 2016: fig. 9.5). Additionally, the generic assignment of material representing C. bohemicus is controversial. Schlosser (1899) originally placed the taxon in the genus Pseudocyon, although more recent contributions have discussed the material as belonging to Amphicyon ( Fejfar & Heizmann 2016, following Kuss 1965) or Cynelos ( Jiangzuo et al. 2018, following Hunt 1998; Hunt & Stepleton 2015). DESCRIPTIONS AND COMPARISONS
In the holotype mandible, CUWM 55, p4, m1, and m2 are present, while the canine, p1-p3, and m3 are represented by alveoli. Judging from the alveoli, C. anubisi n. sp. had a single-rooted p1 and p2, and a small, double rooted p3, with a long diastema between p1-p2, longer than that observed in MNCN 79042 from Gebel Zelten. An alternative possible interpretation is that p 1 in C. anubisi n. sp. is reduced, and the p2 is double-rooted, but this is not the condition observed in MNCN 79042. In this respect, C. anubisi n. sp. clearly differs from the other Moghra amphicyonid CUWM 53, the Gebel Zelten specimen NHM M 82373, Amphicyon giganteus from Arrisdrift ( Morales et al. 2003), and Afrocyon burolleti ( Morales et al. 2010), all of which have a double-rooted p2 and lack diastemata in the premolar series.
The p4 is a typical amphicyonid tooth with the posterior part being broader than the anterior. The tooth is smaller than that of Cynelos macrodon from Kenya ( Adrian et al. 2018), and is similar in size to that of MNCN 79042 from Gebel Zelten, although the p4 of C. anubisi n. sp. is slightly broader. A very low preprotoconulid is indicated by abrasion marks, a feature also present in the Gebel Zelten specimen.The protoconid is the tallest cusp, forms the most voluminous part of the tooth, and also shows strong horizontal abrasion. A strong protoconulid about half the height of the protoconid was present on the distal margin of the tooth, but the protoconulid is abraded in this specimen. A small postprotoconulid is present on the distal end of the postprotocristid, and this feature lacks abrasion marks. Such a postprotoconulid is also present, although smaller, in MNCN 79042. This is in contrast to C. ginsburgi n. comb., where the p4 is relatively shorter compared to m1, with pre- and postprotoconulids present, and C. macrodon, which lacks both a preprotoconulid and postprotoconulid.
The morphology of m1 is preserved in the holotype and is also visible in DPC 14532 ( Morlo et al. 2007). Both specimens are rather slender, with a low metaconid placed slightly distolingual to the protoconid. Both, the m1 of the type specimen, as well as of DPC 14532, are weathered and abraded, so the original heights of the paraconid, protoconid, and hypoconid are unknown. The lingual border of the talonid is occupied by a small hypoconulid and an even smaller entoconid. The m1 of C. anubisi n. sp. is about the same size as in C. macrodon, but C. anubisi n. sp. has a relatively longer paraconid blade, even longer than that observed for C. ginsburgi n. comb. However, C. anubisi n. sp. and C. macrodon share a more angled paraconid blade than C. ginsburgi n. comb., as well as a metaconid that is positioned slightly more anteriorly. The apex of the m1 metaconid in C. ginsburgi n. comb. is tilted slightly distally, but due to abrasion this feature cannot be determined for the m1 of C. macrodon and C. anubisi n. sp.
The m 2 in C. anubisi n. sp. is a massive, elongated tooth in which the protoconid is larger than the metaconid, and the hypoconid approaches the size of the protoconid. Although the tooth is worn and abraded, the original height of the three cusps would have been low. A small entoconid is present on the lingual margin of the tooth. In contrast to Amphicyon , the talonid basin is greatly reduced. As in p4 and m1, there is no development of a cingulid. The very same features are present in an isolated m2 NHM M 82374, assigned to Afrocyon burolleti by Morales et al. (2010: fig. 3) from Gebel Zelten, although the Libyan tooth differs from CUWM 55 in being about 15% smaller. Both the Moghra and Gebel Zelten m2s differ from the holotype of Afrocyon burolleti in being proportionally broader, with a length/breadth ratio of 1.4, while this same figure is about 1.7 in Afrocyon.
The m3 is represented only by its alveolus, although the depth and conical shape of the feature suggests a small but substantial tooth, which would have participated in the grinding function of the molar row. It differs from m3 of Afrocyon in being single-rooted.
The mandibular symphysis in C. anubisi n. sp. extends slightly more posteriorly, past p2, as compared with MNCN 79042 from Gebel Zelten, in which the symphysis ends directly below p2. In addition, the C. anubisi n. sp. mandible has two mental foramina, one located between p1 and p2, and the other situated slightly posterior to p4. In MNCN 79042 the anterior mental foramen is located slightly more anteriorly below p2, and two smaller foramina are present beyond the anterior root of p4.
REMARKS
All African members of the genus Cynelos are united by the presence of diastemata between reduced premolars, a feature that distinguishes Cynelos from the other Early Miocene African amphicyonids, Amphicyon and Afrocyon. The occurrence of a diastema between p2 and p3, and the overall trenchant tooth morphology are features shared among Cynelos from Moghra, the type species of Cynelos (the European C. lemanensis), and African C. euryodon.
Based on the mandible CUWM 55 we erect a new species of Cynelos , C. anubisi n. sp. The m1, DPC 14532/2 ( Morlo et al. 2007) described previously is designed as the paratype. Additionally, a mandibular fragment from Gebel Zelten, MNCN 79042, previously discussed as “ Amphicyon sp.” ( Morales et al. 2010), is also assigned to C. anubisi n. sp., due to a shared similarity in size, a shared low and slender morphology of the mandibular ramus, and the combined presence of a single rooted p1 and p2, a long diastema between p2 and p3, a very short diastema between p3 and p4, the occurrence of pre- and postprotoconulids on p4, and lack of cingulids. In addition, an isolated m2 NHM M 82374 from Gebel Zelten is provisionally referred to C. anubisi n. sp. due to its rectangular outline and small mesiolingual enlargement, which differ both from Afrocyon, and from the m 2 in the mandible NHM M 82373 ( Morales et al. 2010: fig. 2) currently attributed to cf. Amphicyon (see below). We thus assign the tooth to Cynelos anubisi n. sp., while recognizing that the Gebel Zelten specimen is slightly smaller than the m2 of the holotype.
Cynelos anubisi n. sp. differs from similar sized Cynelos in having a longer paraconid blade. However, a thorough comparison between C. anubisi n. sp. and C. macrodon is hampered by the fact that the only teeth available for comparison are p4 and m1. Indeed, among species of Cynelos , C. macrodon is not very well represented. The holotype of C. macrodon is an M1 from Rusinga Island, Kenya ( Savage 1965), and the hypodigm is comprised of only isolated teeth. These include m1 and M1 fragments (Schmidt-Kittler 1987), an additional m1 ( Morales et al. 2016), recent material from Kalodirr, which is a p4, P4, and M1 ( Adrian et al. 2018), and an M2 from Napak-I, assigned to “ Afrocyon sp.” by Morales et al. (2016: figs. 2, 6). Pickford et al. (2003) identified “? Cynelos ” as present in the fauna from Moroto, Uganda, and Leakey et al. (2011) list a few records of C. macrodon from sites in the Turkana Basin. However, except for the material discussed by Adrian et al. (2018), descriptions of C. macrodon are generally lacking. While the m1 of C. macrodon from Rusinga (Schmidt- Kittler 1987, Morales et al. 2016) is about the same size as the m1 of C. anubisi n. sp., the Rusinga specimen lacks the long paraconid blade and shorter protoconid of C. anubisi n. sp. Compared with C. macrodon from Kalodirr ( Adrian et al. 2018), the p4 of C. anubisi n. sp. is smaller and possesses pre- and postprotoconulids. We consider possession of a longer m1 paraconid blade to be an important character distinguishing C. anubisi n. sp. from C. macrodon. Confidence in this feature as a diagnostic character is based on the observation that, among species of Cynelos , members of C. euryodon exhibit variation in a number of ways, including the height of the m1 hypoconid, the size of the antero-labial enlargement, cusp height, and strength of the anterior cingulid in m2, but not in the length of the m1 paraconid blade (pers. obs. MM on KNM without number, KNM RU 2986, KNM RU 4393).
Finally, a lower canine and M2 from the Middle Miocene Muruyur Formation, Kenya, were attributed to C. macrodon by Morales & Pickford (2008), but were subsequently transferred to Myacyon by Morales et al. (2016). The specimen KNM-MY 89 ( Fig. 3 View FIG ) is a left m2 from the same area. This tooth is part of the carnivoran collection described by Schmidt-Kittler (1987), but this particular specimen was not included in his monograph because it was misplaced at the time. The morphology of KNM-MY 89 is much closer to Cynelos than to Myacyon (see Morales et al. 2010: fig. 3, 6) because it is nearly identical to the m2 of CUWM 55 and to NHM M 82374 from Gebel Zelten. Due to this morphological resemblance we refer the lower canine, M2, and m2 from the Muruyur Formation to Cynelos cf. macrodon, thereby re-instating the initial view of Morales & Pickford (2008). These specimens represent the only record of a large middle Miocene Cynelos in Africa.
Morales et al. (2010) referred the Moghra specimens described by Morlo et al. (2007) as “ Cynelos sp. nov.” to the species “ Ysengrinia ” ginsburgi from Arrisdrift ( Morales et al. 1998: fig. 4; Morales et al. 2003; Werdelin & Peigné 2010). Due to the presence of a large M2, Morales et al. (2016) excluded the taxon from Ysengrinia and transferred it to Afrocyon. However, the Arrisdrift species differs from Afrocyon burolleti and resembles C. anubisi n. sp. in possessing a much more trenchant dentition with narrower m1 and m2 talonids, and in having a diastema between p2 and p3 ( Morales et al. 1998, 2003). We thus corroborate the exclusion of the species from Ysengrinia, but transfer it from Afrocyon to Cynelos as C. ginsburgi n. comb. Cynelos anubisi n. sp. and C. macrodon differ from C. ginsburgi n. comb. in having the m1 metaconid located slightly more anteriorly, and C. anubisi n. sp. further differs from C. ginsburgi n. comb. in having p4 and m2 larger relative to m1. Cynelos ginsburgi n. comb. differs from C. macrodon in having the P4 protocone less reduced, M1 less triangular, and pre- and postprotoconulids present on p4 (see Morales et al. 1998). Cynelos ginsburgi n. comb. also has a shortened M1 talon (see Adrian et al. 2018 for C. macrodon and Morales et al. 2016 for C. ginsburgi n. comb.) relative to C. macrodon, but this abbreviated talon occurs to a much lesser degree in C. ginsburgi n. comb. than it does in Amphicyon giganteus from Moghra.
Genus Amphicyon Lartet in Michelin, 1836 (see Peigné 2012)

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.