
Callogobius pilosimentum, Delventhal, Naomi R., Mooi, Randall D., Bogorodsky, Sergey V. & Mal, Ahmad O., 2016
|
publication ID |
https://doi.org/10.11646/zootaxa.4179.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:A456F585-0F3A-4F98-99CC-795A1A3603C4 |
|
DOI |
https://doi.org/10.5281/zenodo.5679403 |
|
persistent identifier |
https://treatment.plazi.org/id/03B187A8-CD13-FE0C-4C9F-FF5E58D3FA25 |
|
treatment provided by |
Plazi |
|
scientific name |
Callogobius pilosimentum |
| status |
sp. nov. |
Callogobius pilosimentum View in CoL sp. nov.
Hairy-chinned Flapheaded Goby ( Figures 1–2 View FIGURE 1 View FIGURE 2 , Table 1 View TABLE 1 )
Drombus irrasus (non Smith)—Goren 1979: 36.
Callogobius irrasus View in CoL (non Smith)— Goren 1980: 213; Dor 1984: 241.
Callogobius maculipinnis View in CoL (non Fowler)— Goren et al. 1991: 299; Goren & Dor 1994: 53; Golani & Bogorodsky 2010: 46.
Material examined. All specimens collected from the Red Sea.
Holotype. SMF 35756 (KAU12-0224), female, 36.4 mm SL, Saudi Arabia, Farasan Island , N16°43.083' E42°03.934', isolated coral patch of lagoon, 3–5 m, S.V. Bogorodsky & T.J. Alpermann, 20 February 2012 ( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 A). GoogleMaps
Paratypes ( 17 specimens, 25.5–67.0 mm SL). Egypt: BPBM 21518 About BPBM , male?, 41.5 mm SL, El Hameira , coral knoll in 12 m, J.E. Randall & O. Gon, 25 April 1977 ; Saudi Arabia: KAUMM 373 , male, 47.1 mm SL, Maqna , N28°26'13.40" E034°45'47.80", steep slope, 7 m, S.V. Bogorodsky, 13 April 2011 GoogleMaps ; KAUMM 374 , female, 28.3 mm SL, Jeddah , Obhur (Sharm Obhur), N21°42'33.12" E39°05'48.26", base of rocky reef, 2 m, S.V. Bogorodsky, 19 April 2011 GoogleMaps ; KAUMM 375 (KAU12-0220), male, 25.5 mm SL, collected with the holotype; KAUMM 376 , male, 37.0 mm SL, Farasan Island, N16°43.083' E42°03.934', isolated coral patch of lagoon, 3–5 m, S.V. Bogorodsky & T.J. Alpermann, 20 February 2012 GoogleMaps ; KAUMM 377 (KAU13-378), male, 28.5 mm SL, 30 km south of Al Wajh , N26°03'30.36" E36°38'34.98", fringing reef, 8 m, S.V. Bogorodsky & T.J. Alpermann, 14 June 2013 GoogleMaps ; SMF 35757, 2 males, 29.7 mm SL (C&S) & 47.3 mm SL, Farasan Island, N16°54' 93.30" E41°50'76.10", lagoon, isolated small coral patch, 1 m, S.V. Bogorodsky, 0 1 April 2011 GoogleMaps ; SMF 35758 (KAU12-0218), male, 38.4 mm SL, collected with the holotype; SMF 35759 (KAU12-0538), female, 35.8 mm SL, Farasan Island, N16°43.083' E42°03.934', isolated coral patch of lagoon, 3–5 m, S.V. Bogorodsky & T.J. Alpermann, 28 February 2012 GoogleMaps ; SMF 35760, male, 67.0 mm SL and female, 55.8 mm SL ( Fig. 2 View FIGURE 2 C), Jeddah , Obhur (Sharm Obhur), N21°42'32.28" E39°05'47.16", steep slope with patches of corals and small sandy flats, 14–16 m, S.V. Bogorodsky, 0 1 July 2013 GoogleMaps ; Sudan: BMNH 1978.9.8.12-16, 5 specimens ( 4 females with 1 C&S, 1 male, 44.5–61.9 mm SL), Suakin, coral crevices in seawall, Manihine collections, 5 December 1950 .
Other material. BMNH 1978.9.8.17-26, 10 specimens ( 6 females, 4 males, 36.9–70.7 mm SL), Sudan, Suakin Archipelago , around seawall and from crevices, Manihine collections, 12 January 1951.
Tentative identification: USNM 296956 About USNM , female, 44.3 mm SL, Red Sea, Egypt, just north of Ras Burqa , V.G. Springer et al., 23 July 1969.
Diagnosis. Callogobius pilosimentum is distinguished from all other known Callogobius species by the following combination of characters: interorbital canal normally containing pores B’, C, D, E, F, G and H’; preopercular canal containing pores M’, N, and O’; temporal canal containing pores K’ and L’; scales in lateral series 21–25 (usually 24); normally four transverse mandibular papillae rows (Row 16) on each side.
Description. Holotype values indicated by an asterisk. Parentheses enclose number of type specimens with the particular value, counts made on both sides when applicable or possible. Dorsal fin VI + I,9(17*), VI + I,10(1); anal fin I,6(1), I,7(17*); pectoral-fin rays 15(2), 16(11), 17(15*), 18(5); pelvic-fin rays I,5(36*); segmented caudalfin rays 9 + 8(1), i8 + 8(4*), i8 + 7i (12), ii7 + 8(1); procurrent rays 4 + 4(2), 5 + 4(15*), 5 + 5(1); scales in lateral series 21(1), 22(3*), 23(4), 24(20*), 25(5); predorsal scales 6(5), 7(13*); transverse scales 8(3), 9(13*), 10(2).
See Table 1 View TABLE 1 for selected morphometrics.
Body moderately robust for the genus ( Fig. 2 View FIGURE 2 ). Head depressed, broader than deep. Snout obtuse. Mouth slightly oblique, forming an angle of about 45º with body axis; lower jaw slightly beyond tip of upper jaw, posterior end of jaws at or just before vertical through anterior margin of orbit. Anterior nostril moderately long, slender tube, reaching halfway to anterior outer edge of upper lip; posterior nostril very short, upright tube. Eye moderately large; upper margin of orbit slightly elevated above profile of head. Interorbital narrow. Gill opening ending ventrally at lower edge of pectoral-fin base. Tongue broad, tip slightly bilobed. Urogenital papilla long, slender and darkly pigmented in males; broad and darkly pigmented in females, no lateral flaps. Vertebral count (based on two cleared and stained specimens) 10 precaudal + 16 caudal.
Dentition. Teeth in jaws conical and slender in rows; outer teeth slightly larger than inner teeth. Outer teeth present on anterior two-fifths of toothed portion of lower jaw; inner teeth of lower jaw tightly packed anteriorly, about 6 irregular rows merging to single row posteriorly. Outer teeth of upper jaw extend over entire toothed portion of upper jaw, about 5 irregular rows of inner teeth merging to single row parallel to row of outer teeth.
Fins. First dorsal-fin base short; anterior fin membranes incised, first three interspinous membranes incised with spine tips free up to one-fourth their length, occasionally more for second and third spine (particularly in males that have longer, filamentous fin spines), posterior interspinous membrane only slightly incised; second spine longest. Second dorsal-fin base about twice that of first dorsal; segmented rays branched, penultimate ray longest. Anal-fin base short; segmented rays branched, penultimate ray longest. Pectoral-fin rays branched except for dorsalmost one or two rays; fin reaching to level of second or third anal-fin ray. Pelvic fins fully united with membrane over entire length of medial rays; fourth segmented pelvic-fin ray barely shorter or subequal to fifth ray, fifth ray almost reaching to anus; all segmented pelvic-fin rays branched. Pelvic frenum present and well developed. Caudal fin rounded, length <40% SL.
Squamation. All scales large and deciduous, with distinctly outlined centres. Scales cycloid on head and anterior half of body; cycloid scales present on nape to interorbital, in spaces between papillae rows on cheeks, preoperculum and operculum, on lateral side of pectoral-fin bases, prepelvic and predorsal areas, and belly. Scales ctenoid on posterior half of body from mid-flank region below first to third spines of first dorsal fin. Ctenii numerous, long, slender and pointed; no elongate ctenii on caudal peduncle. Most specimens with single large cycloid scale in centre of pelvic disc (visible under pelvic frenum).
Cephalic sensory systems. Pores present ( Fig. 1 View FIGURE 1 ). Interorbital canal with pair of posterior nasal pores (pore B), anterior interorbital pore (pore C), posterior interorbital pore (pore D), pair of supraotic pores (pore E), pair of anterior otic pores (pore F), pair of posterior otic pores (pore G), and pair of intertemporal pores (pore H); preopercular canal with three preopercular pores (pores M, N and O); each temporal canal with anterior and posterior temporal pore (pores K and L, respectively).
Papillae row configuration ( Fig. 1 View FIGURE 1 ): Postnasal rows (Row 2) long and joined across midline (16*) or overlapping (1). Anterior suborbital row (Row 9) short to moderately long, not reaching eye (34*). Mid suborbital row (Row 10) short to moderately long, not reaching eye (34*). Posterior suborbital rows (Row 11) long and overlapping (32*) or rarely short or irregular (2). Longitudinal maxillary row (Row 14) rarely irregular (1), normally continuous and extending posteriorly to below and beyond (33*) transverse cheek row (Row 13), latter short (31*) or rarely irregular (1). Longitudinal mandibular row (Row 15) continuous (21*) or irregular (3). With 4(31*), 5(2), or 2(1) transverse mandibular rows (Row 16) on each side of lower jaw, all but one specimen with 4 on at least one side; 15 of 17 type specimens with 4 on both sides. One specimen with 5 on one side and 4 on the other (SMF 35760, 67 mm SL), and one with 5 on one side and 2 on the other (SMF 35759, 35.8 mm SL); latter exhibits irregular patterns in several other papillae rows (Rows 13, 14, 15, 17, 20, 21). Postorbital rows (Row 17) short (less than two-thirds of distance from the dorsal mid-line to bony edge of cranium) (8) or medium (more than two-thirds of distance from dorsal mid-line to bony edge of the cranium) (24*), rarely irregular (1). Preopercular row (Row 20) continuous with the transverse opercular row (Row 21) (25*) or separate (5).
Color in life ( Fig. 2 View FIGURE 2 ). General color pattern brown to dark brown with paler patches ( Fig. 2 View FIGURE 2 A), or pale grayishbrown with up to five dark brown mottled vertical bars on body and dark brown blotchy patterns with varying contrast on head. Body bars, when visible (e.g. Figs. 2 View FIGURE 2 B & C, but not Fig. 2 View FIGURE 2 A), appear as follows: irregular bar may extend dorsally over operculum and pectoral-fin base, slightly indented at first dorsal-fin spine; wide bar extends below first dorsal fin; two wide, slightly oblique bars extend below mid and posterior portions of second dorsal fin; narrow bar encircles posterior caudal peduncle, edge of hypural plate and proximal regions of the caudal fin. Underside of head light brown or dusky, papillae rows may show great color contrast. First dorsal fin with dark brown basal spot centered on fourth ray and continuous with dark body bar; may exhibit yellow markings on distal margin. First and second dorsal fins dark brown with irregular, oblique, pale bands or rows of spots, some individuals with irregular dark and pale pattern; caudal fin dark brown usually with pale spots forming irregular bars. Pelvic and anal fins dusky or dark brown with or without paler blotches. Pectoral fin with dark irregular bands or rows of spots and a dark medial mark at base. Urogenital papilla dark.
Color in preservative. As in life, although older specimens can be orange-brown with little pattern on body. Most museum specimens damaged (species with fragile skin and deciduous scales) and body color pattern often reduced to light brown or grey with dark outlines of scale pockets.
Etymology. The species name is derived from the Latin pilosus meaning “hairy” and mentum meaning “chin”, referring to the extra rows of papillae on the chin. Specific epithet to be treated as a noun in apposition. Suggested common name: hairy-chinned flapheaded goby.
Distribution and habitat. Restricted to the Red Sea. It is usually collected in shelters or close to shelters on small sand flats (about 0.5–1 m across) of steep slopes, sometimes at the base of coral reefs, in closed areas in bays and lagoons, at depths of 1– 20 m.
Remarks. Goren (1980) listed Drombus irrasus Smith in the Red Sea as a first record. This species is considered a synonym of Callogobius maculipinnis by most authors (e.g. McKinney & Lachner 1984, Goren et al. 1991); Eschmeyer & Fricke (2016) incorrectly attributed Delventhal & Mooi (2013) as recognizing Smith’s species when the type was listed as comparative material. Although we have not examined Goren’s (1980) C. irrasus material, we suspect that these specimens will belong to C. pilosimentum . Two lots of specimens that we identify as C. pilosimentum (BMNH 1978.9.8.12-16 and BMNH 1978.9.8.17-26) were examined by Goren (1980) and identified as C. irrasus .
Callogobius pilosimentum View in CoL belongs to a “ maculipinnis View in CoL species complex” including the following nominal species: C. bauchotae Goren 1979 View in CoL , C. irrasus ( Smith 1959) View in CoL , C. kuderi ( Herre 1943) View in CoL , C. maculipinnis ( Fowler 1918) View in CoL , C. nigromarginatus Chen & Shao 2000 View in CoL , C. shunkan Takagi 1957 , C. snelliusi Koumans 1953 View in CoL , and C. vanclevei ( Herre 1950) View in CoL . Species of this complex are characterized by having the temporal canal with pores K and L (absent in all other Callogobius View in CoL ) and normally four or fewer transverse mandibular rows on each side (all other Callogobius View in CoL normally have more than 10). McKinney & Lachner (1984) synonymised C. irrasus View in CoL , C. kuderi View in CoL , C. shunkan , C. snelliusi View in CoL , and C. vanclevei View in CoL with C. maculipinnis View in CoL ; they did not examine C. bauchotae View in CoL (or C. nigromarginatus View in CoL as it was described after their work). However, their synonymies should be considered questionable because all included species are poorly defined. Based on the first author’s examination of the types, we are confident that several, or even most, of these species will prove to be valid after complete investigation. Despite this, C. pilosimentum View in CoL can readily be distinguished from the other members of this complex, all of which normally have three transverse mandibular rows on each side. We previously examined the types of all the above species in the maculipinnis View in CoL complex except C. nigromarginatus View in CoL . They all have three sets of transverse mandibular rows, except C. shunkan which has extra irregular rows and is easily differentiated by other characters (Y. Ikeda pers. comm.). These observations of the types were recently confirmed by Y. Ikeda ( C. bauchotae View in CoL , C. snelliusi View in CoL , and C. shunkan ), M. Sabaj Pérez ( C. maculipinnis View in CoL ) and D. Catania ( C. kuderi View in CoL ). L. Parenti re-examined the holotype of C. vanclevei View in CoL , a distorted specimen that appears to have no fourth transverse mandibular row on either side. The holotype of C. nigromarginatus View in CoL was unavailable, but the illustration and text indicate that three transverse mandibular rows are present ( Chen & Shao 2000).
We found only a single Red Sea specimen (USNM 296956, female, 44.3 mm SL) that has three transverse mandibular rows on each side. We tentatively identify it as Callogobius pilosimentum , but we have not designated it as a paratype. We presume that this is simply unusual variation in a species that normally has four on each side. To evaluate variation in transverse mandibular papillae rows (Row 16, Fig. 1 View FIGURE 1 ) in specimens outside of the Red Sea, we examined 174 adult specimens identified in collections as C. maculipinnis ranging across the Indian Ocean eastward to Mangareva (Gambier Islands, French Polynesia) in the Pacific Ocean (listed in Comparative material below).
Of the 174 specimens examined from outside of the Red Sea, only three (<1.7%) exhibited the condition of C. pilosimentum of having four transverse mandibular papillae rows on both sides of the jaw. Even among these, only one (WAM P27935-024) exhibited a normal, sequential four-row anatomy, and this specimen belongs to an Australian ‘population’ that has distinctly higher meristic counts than other members of the complex (D. Hoese pers. comm.). The other two specimens (USNM 332226 and ZRC 40669) have unusual morphologies where the fourth transverse mandibular row on one side follows a free papilla and/or the first row is a branch of the intermandibular row. An additional 11 specimens had four rows on only one side and three or fewer on the other (6.3% of all specimens). From localities with the largest representation, an Indonesian lot (USNM 241882) had three of 32 specimens having four transverse mandibular papillae rows on one side only and of 42 specimens in 11 lots from Fiji only two specimens had four such rows on one side. This is in contrast to the condition in C. pilosimentum in the Red Sea where all specimens exhibited four or more transverse mandibular rows on each side except one that had five on one side and two on the other, and one that had three on each side (USNM 296956).
In Callogobius pilosimentum , the anterior first dorsal-fin spines are usually free of membrane for one-fourth or more of the spine length, particularly the second spine. Although clearly evident on some live specimens ( Fig. 2 View FIGURE 2 ), this can be difficult to assess on preserved specimens. This feature is common to most, if not all, members of the maculipinnis species complex so is not diagnostic, but is potentially of value to distinguish live or particularly well-preserved specimens from Red Sea congeners.
Comparative material. Callogobius maculipinnis (listed west to east): Comoros—ROM 92691 (4); Oman— ROM 39895 (6); Chagos—ROM 55107 (4); Sri Lanka—USNM 220035 (5); Cocos-Keeling—WAM P29928-022 (4); Western Australia—WAM 27935-024 (3); Thailand—ROM 58036 (1), 68041 (4); Indonesia—USNM 241882 (32), WAM P33093-002 (2); Singapore—ZRC 40669 (1); Philippines—ROM 53339 (3), USNM 297102 (2); Taiwan—BPBM 23242 (1), USNM 298439 (8); Palau—BPBM 37767 (2), ROM 75955 (1); 76134 (1); Papua New Guinea—BPBM 32638 (1), 32674 (2), USNM 297051 (2); Australia—MPM 48365 (1), ROM 38903 (1), USNM 297048 (3); One Tree Island—BPBM 14432 (4); Lord Howe Island—BPBM 14862 (1); Coral Sea—BPBM 33517 (1), 33629 (3); New Caledonia—BPBM 34270 (1), ROM 64155 (1), 64160 (2), 64416 (3); Vanuatu—BPBM 5765 (1), MPM 32132 (1), 46721 (1); Marshall Islands—BPBM 8296 (1), 17739 (1), 22349 (1); Fiji—BPBM 39866 (1), 40082 (1), ROM 57700 (5), 57701 (1), 57703 (3), 57705 (6), 57707 (2), 57708 (1), 57709 (2), 57710 (1), USNM 332226 (19); Tonga—BPBM 38115 (3), USNM 339828 (7); Tahiti—BPBM 8315 (2); Rapa—BPBM 17317 (1); Mangareva—BPBM 13595 (3).
Comparisons to other described Callogobius species are based on our data from the available holotypes listed in Delventhal & Mooi (2013).
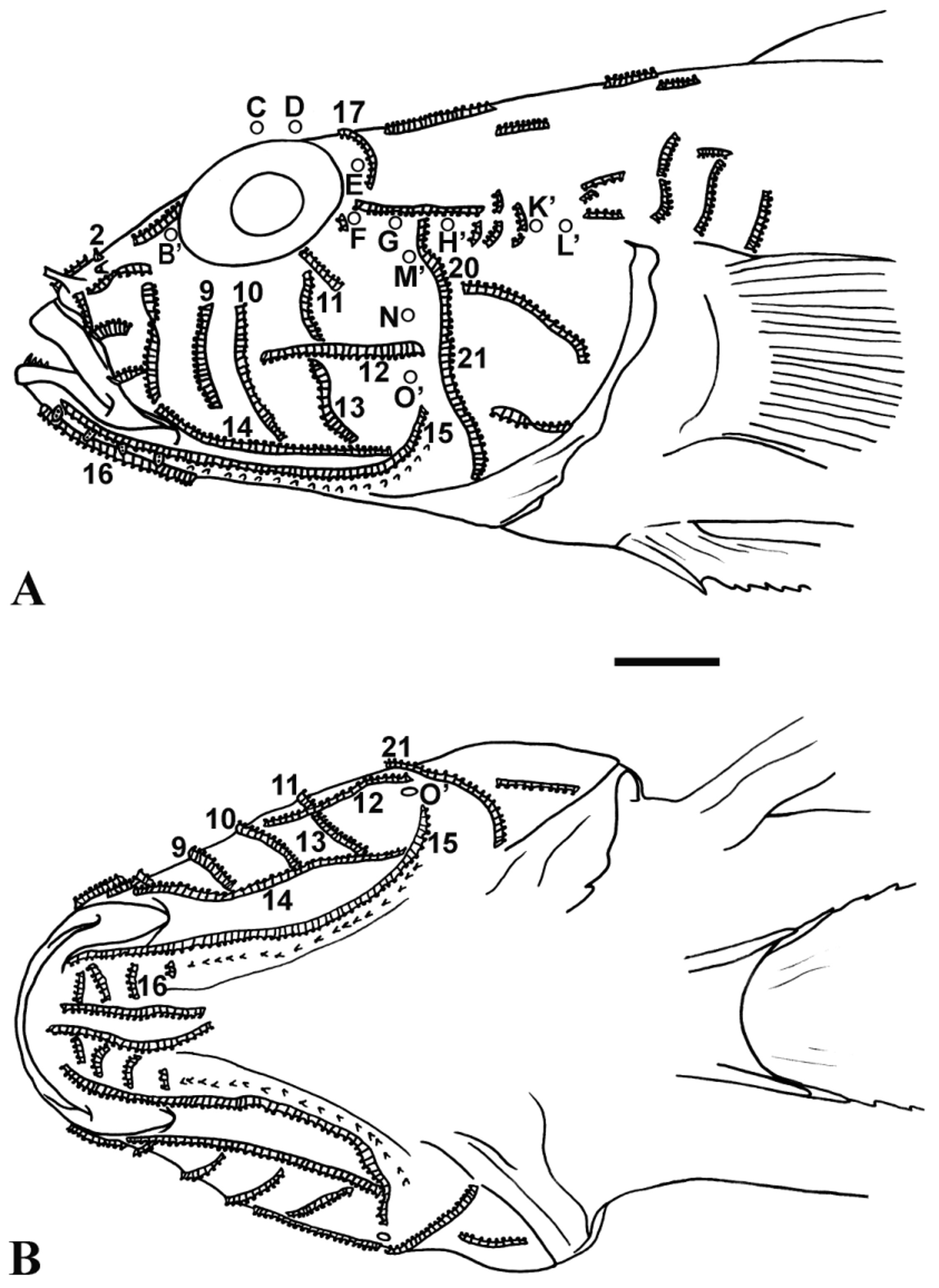

TABLE 1. Selected measurements of the holotype and paratypes of C. pilosimentum sp. nov.; standard length is provided in mm, values for other morphometrics are as percentage of standard length (SL) or head length (HL), as indicated. For paratypes, the range is followed by an average value for all types in parentheses. The two largest specimens (63 and 67 mm SL) were males (M), the next 3 largest specimens (55.8, 50.3, 49.3 mm SL) were females (F).
| Holotype | Paratypes | |
|---|---|---|
| Gender | F | 9M,8F |
| Standard Length (mm) | 36.4 | 25.5–67.0 (42.8) |
| Head length (% of SL) | 33.0 | 29.1–34.1 (32.3) |
| Head depth (% of HL) | 59.2 | 50.6–64.6 (58.8) |
| Head width (% of HL) | 73.3 | 64.6–75.8 (70.2) |
| Interorbital width (% of HL) | 5.4 | 4.1–7.8 (5.7) |
| Predorsal fin distance (% of SL) | 38.5 | 34.3–40.4 (37.0) |
| Preanal fin distance (% of SL) | 59.6 | 57.8–62.8 (60.3) |
| Prepelvic fin distance (% of SL) | 33.0 | 29.6–35.8 (32.8) |
| Pectoral-fin length (% of SL) | 31.3 | 29.7–35.4 (32.6) |
| Pelvic-fin length (% of SL) | 23.9 | 22.0–28.1 (24.9) |
| Caudal-fin length (% of SL) | 32.4 | 26.4–37.3 (31.9) |
| Caudal peduncle depth (% of SL) | 13.7 | 12.3–15.5 (13.8) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Callogobius pilosimentum
| Delventhal, Naomi R., Mooi, Randall D., Bogorodsky, Sergey V. & Mal, Ahmad O. 2016 |
Callogobius maculipinnis
| Golani 2010: 46 |
| Goren 1991: 299 |
Callogobius irrasus
| Goren 1980: 213 |