
Pleuroceros blanfordi, (LYDEKKER, 1884)
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2009.00597.x |
|
persistent identifier |
https://treatment.plazi.org/id/03B187BE-B275-FFB7-FF0C-DD9EBEA10F9D |
|
treatment provided by |
Valdenar |
|
scientific name |
Pleuroceros blanfordi |
| status |
|
PLEUROCEROS BLANFORDI ( LYDEKKER, 1884)
COMB. NOV. ( FIGS 4–7 View Figure 4 View Figure 5 View Figure 6 , 11A View Figure 11 , 12A View Figure 12 )
Rhinoceros palaeindicus Lydekker, 1881: 44–45 ; pl. 6, fig. 1
Aceratherium blanfordi sp. nov., nobis Lydekker, 1884: 2–11, text-fig. 2; pl. 2, figs 1–3
Aceratherium blanfordi var. majus Lydekker, 1884: 10 ; pl. 1, 1–2
Rhinoceros blanfordi var. majus Lydekker, 1886: 154
Aceratherium blanfordi Lyd. Pilgrim, 1908: 149
Aceratherium blanfordi var. majus Pilgrim, 1910: 66
Teleoceras blanfordi Lydekker Pilgrim, 1912: 3 , 30–32, pl. 7, figs 4–7
Chilotherium blanfordi Ringström, 1924: 75
Chilotherium blanfordi Matthew, 1929: 508
Chilotherium blandfordi Forster-Cooper, 1934: 589– 594 ; text-fig. 9, 12C; pl. 67, figs 34–38
« Dicerorhinus View in CoL » cf. abeli (partim) Welcomme et al., 1997: 532, 535
? « Dicerorhinus » cf. abeli (partim) Welcomme et al., 1997: 534, 535, 536
Aprotodon blanfordi Welcomme & Ginsburg, 1997: 1001 , table
Pleuroceros blanfordi Lindsay et al., 2005 : table 1
‘ Aprotodon ’ blanfordi Métais et al. 2009: 163 , 164; table 2, fig. 5
Emended diagnosis: Differs from P. pleuroceros by its larger size (c. 15%), the presence of a posterior horizontal groove on the processus zygomaticus of the squamosal, the absence of a sagittal lingual groove on the corpus mandibulare, a shortened premolar series, higher tooth crowns, the abundance of coronary cement, a weaker labial cingulum, a multiple crochet always present, an unconstricted metaloph, a continuous lingual cingulum, and a thick lingual bridge on upper premolars, a transverse metaloph and a reduced protocone on P2, and the usually constricted protocone on P3–4, the absence of a crista on P3, the unconstricted metaloph on P4, the usual presence of a lingual cingulum (occasional in P. pleuroceros ) on upper molars, a deep protocone constriction and the presence of a metacone fold on M1–2, a strong mesostyle on M2, a constricted metaconid on lower deciduous teeth, the absence of a posterior McIII-facet on McII, the absence of a fibula-facet on the calcaneus, and the concave proximal border of MtIII.
Lectotype: Right P4–M2 series unearthed in Gandoï, Bugti Hills (IMC C. 268) and figured by Lydekker (1884: pl. 1, fig. 1), proposed as a lectotype by Pilgrim (1912: 31).
Type locality: Gandoï, Bugti Hills, Pakistan (Early Miocene?).
Stratigraphical range: Chitarwata Fm. (Bugti and Zinda Pir areas) and base of the Vihowa Fm. (Bugti area). Early Miocene (c. 23–18.5 Myr; Lindsay et al., 2005; Métais et al., 2009).
Geographical range: Bugti and Zinda Pir area, Sulaiman Lobe, Balochistan, Pakistan.
Referred material
Old collections: ‘Siwaliks of the Punjab’ [?Early Miocene]. Left maxilla with P4–M2 IMC-without number; Gandoi [?Early Miocene, Bugti Hills]. Right mandible with p3–m1 and m2–3 salient (IMC C. 271); left M2 (IMC C. 259); right M1 or M2, worn (IMC C. 262); part of a right juvenile mandible, with d3 (IMC C. 267). Dera Bugti [?Early Miocene]. Part of left maxilla with M1–3 (IMC C. 268). ‘Gaj of the Bugti Hills’ [?Early Miocene]. M1–3 (IMC C. 266). ‘Near Dera Bugti’ [locality and age unknown]. Germ of a left P2 (NHM M 15335 View Materials ); left P2 and P3 from the same individual (NHM M without number [w.n.]); left P3 and P4 from the same individual (NHM M w.n.); fragment of a right maxilla with D2–4 and M1 (NHM M 15367) and germ of P3 extracted from the maxilla (NHM M 15368); right P2 (NHM M 15337); right P3 (NHM M 15338); left P3 and P4 from the same individual (NHM M 15333); right P3 and P4 from the same individual (NHM M 15366); palate with left P2–M3 and right M1–2 (NHM M 15365).
New material
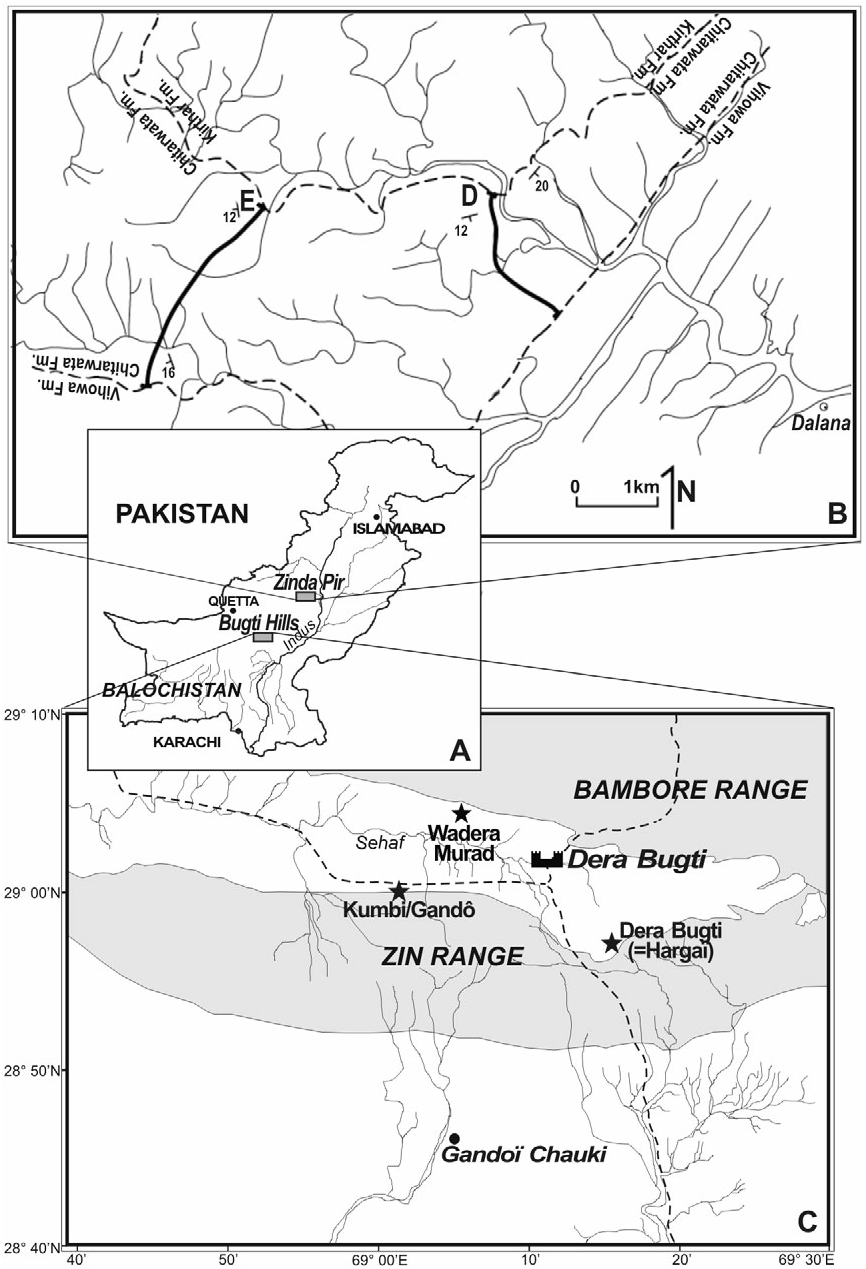
Bugti Hills ( Figs 1 View Figure 1 , 2)
Kumbi 4a (Level 4, earliest Miocene). Mandible from a young adult, in three parts: left corpus with p4–m1, right corpus with p4–m3 and symphysis bearing left i1 and left and right i2, all of them broken (MHNT Pak 1038); fragment of a left mandible with p2 (broken) and p3 (MHNT Pak 1037); fragment of an toothless symphysis (MHNT Pak 1073); fragment of a right mandible with m3 (MHNT Pak 1068); fragment of a left maxilla with P2–P4, M1 without ectoloph and fragments of i2 and right p2 from the same individual (MHNT Pak 1031); right P3 (MHNT Pak 1024); right P3 (MHNT Pak 1059); left P4 (MHNT Pak 1058); right P4 d (MHNT Pak 1046); right P4 without ectoloph (MHNT Pak 1050); fragment of a left maxilla with M1–2 (MHNT Pak 1022), from the same individual as a right M2 (MHNT Pak 1019); right M1 (MHNT Pak 1061); fragment of a right M1 on a maxilla (MHNT Pak 1035); lingual fragment of a right M1 (MHNT Pak 1064); left crushed M2 (MHNT Pak 1045); lingual fragment of a right M2 (MHNT Pak 1027); left M3 (MHNT Pak 1013); right M3 (MHNT Pak 1014); rostral fragment of a left i2 (MHNT Pak 1021); lingual fragment of a right p3 (MHNT Pak w. n.); posterior fragment of a right m3 (MHNT Pak 1020); fragment of a humeral distal end (MHNT Pak 1085); fragment of a left humeral distal end (Pal 1198); proximal end of a left radius (MHNT Pak 1088); proximal end of a right radius (MHNT Pak 1087); proximal end of a right radius (MHNT Pak 1089); distal end of a left radius (MHNT Pak 1091); distal end of a right radius (MHNT Pak 1090); distal end of a right radius (MHNT Pak 1206); right scaphoid (MHNT Pak 1098); right semilunate (MHNT Pak 1101); left magnum (MHNT Pak 1110); right magnum (MHNT Pak 1093); left unciform (MHNT Pak 1112); right unciform (MHNT Pak 1094); right unciform (MHNT Pak 1113); right unciform without posterior tuberosity (MHNT Pak 1114); proximal end of a left McIII (MHNT Pak 1118); proximal end of a left McIII (MHNT Pak 1119); proximal end of a left McIII (MHNT Pak 1120); left McIII (MHNT Pak 1121); right Mc III without distal end (MHNT Pak 1117); distal end of a right Mc III (MHNT Pak 1193); left patella (MHNT Pak 1131); left patella (MHNT Pak 1132); distal end of a left tibia (MHNT Pak 1124); distal end of a left tibia (MHNT Pak 1126); distal end of a right tibia (MHNT Pak 1127); distal end of a right tibia (MHNT Pak 1128); distal end of a right fibula (MHNT Pak 1129); left astragalus (MHNT Pak 1137); left astragalus (MHNT Pak 1138); left astragalus (MHNT Pak 1139); medial fragment of a left astragalus (MHNT Pak 1143); right astragalus (MHNT Pak 1140); right astragalus (MHNT Pak 1141); right tuber calcanei (MHNT Pak 1104); right calcaneus (MHNT Pak 1150; right calcaneus (MHNT Pak 1151); right calcaneus (MHNT Pak 1152); left navicular (MHNT Pak 1154); posterior fragment of a left navicular (MHNT Pak 1156); right cuboid (MHNT Pak 1158); right cuboid (MHNT Pak 1159); right ectocuneiform (MHNT Pak 1160); fragment of a left ectocuneiform (MHNT Pak 1161); right mesocuneiform (MHNT Pak 1590); proximal end of a left MtII (MHNT Pak 1163); distal end of a left MtII (MHNT Pak 1191); right MtII (MHNT Pak 1162); proximal end of a right MtIII (MHNT Pak 1096); distal end of a right MtIII (MHNT Pak 1192); distal end of a right MtIII (MHNT Pak 1194); right MtIII without proximal end (MHNT Pak 1195); distal end of a right MtIV (MHNT Pak 1097); right Mt IV (MHNT Pak 1165); proximal end of a right MtIV (MHNT Pak 1166); proximal end of a right MtIV (MHNT Pak 1167).
Kumbi 4b (Level 4, earliest Miocene). Fragment of an eroded right mandible with erupting m3 (MHNT Pak 772); right juvenile mandible with d2–4, m 1 in the dentary and alveolus of d1 (MHNT Pak 784); left P2 (MHNT Pak 751); right P2 (MHNT Pak 844); left P3 without an ectoloph (MHNT Pak 842); right P3 without an ectoloph (MHNT Pak 758); fragment of a left M2 (MHNT Pak 760); fragment of a protoloph of left M3 (MHNT Pak 761); left M3 without a protoloph (MHNT Pak 763); right M3 (MHNT Pak 918); slightly worn left m3 (MHNT Pak 917); fragment of a left m3 (MHNT Pak 774); right scaphoid (MHNT Pak 785), fragment of a right semilunate (MHNT Pak 786) and right trapezoid (MHNT Pak 787) probably from the same individual; distal end of a left McII (MHNT Pak 789); distal end of a left MtII (MHNT Pak 790).
Kumbi 4c (Level 4, earliest Miocene). Right P2 (MHNT Pak 844); left P3 without an ectoloph (MHNT Pak 845); left M2 (MHNT Pak w. n.); left damaged patella (MHNT Pak 86); distal end of a right tibia (MHNT Pak 71).
Kumbi 4d (Level 4, earliest Miocene). Fragmentary skull with left and right squamosals (processus zygomaticus), left postglenoid apophysis, occipital, left and right maxilla bearing P3, right M1 and the alveoli of right P1–2, fragments of right M2–3 and undetermined fragments (MHNT Pak 46).
Kumbi 4f (Level 4, earliest Miocene). fragment of a right M3 (MHNT Pak 1676); proximal end of a left McII (MHNT Pak 1733); right patella (MHNT Pak 1687).
Gandô 4 (Level 4, earliest Miocene). Fragment of a right M3 (MHNT Pak 1864); left p3 (MHNT Pak 1862).
Dera Bugti 4 (Level 4, earliest Miocene). Fragment of a left P4 (MHNT Pak 1967); right P4 (MHNT Pak 1964).
Dera Bugti 5 (Level 5, Early Miocene). Posterolingual fragment of a left M2 (MHNT Pak 1258).
Dera Bugti 6 (Level 6, Early Miocene). left M 2 g (MHNT Pak 1012a); lingual fragment of a right M3 (MHNT Pak 1444); lingual fragment of a left m1 (MHNT Pak 2215); damaged right astragalus (MHNT Pak 2235).
Wadera Murad (Early Miocene, northern side of the Dera Bugti syncline). Fragment of a left P4 (MHNT Pak 2458).
Zinda Pir Dome ( Fig. 1 View Figure 1 )
Z149 (earliest Miocene). Left astragalus (PMNH Z2043); Z139 (earliest Miocene). Right fragmentary astragalus (PMNH Z2047). Z135 (earliest Miocene). Right p2 (PMNH Z2070).
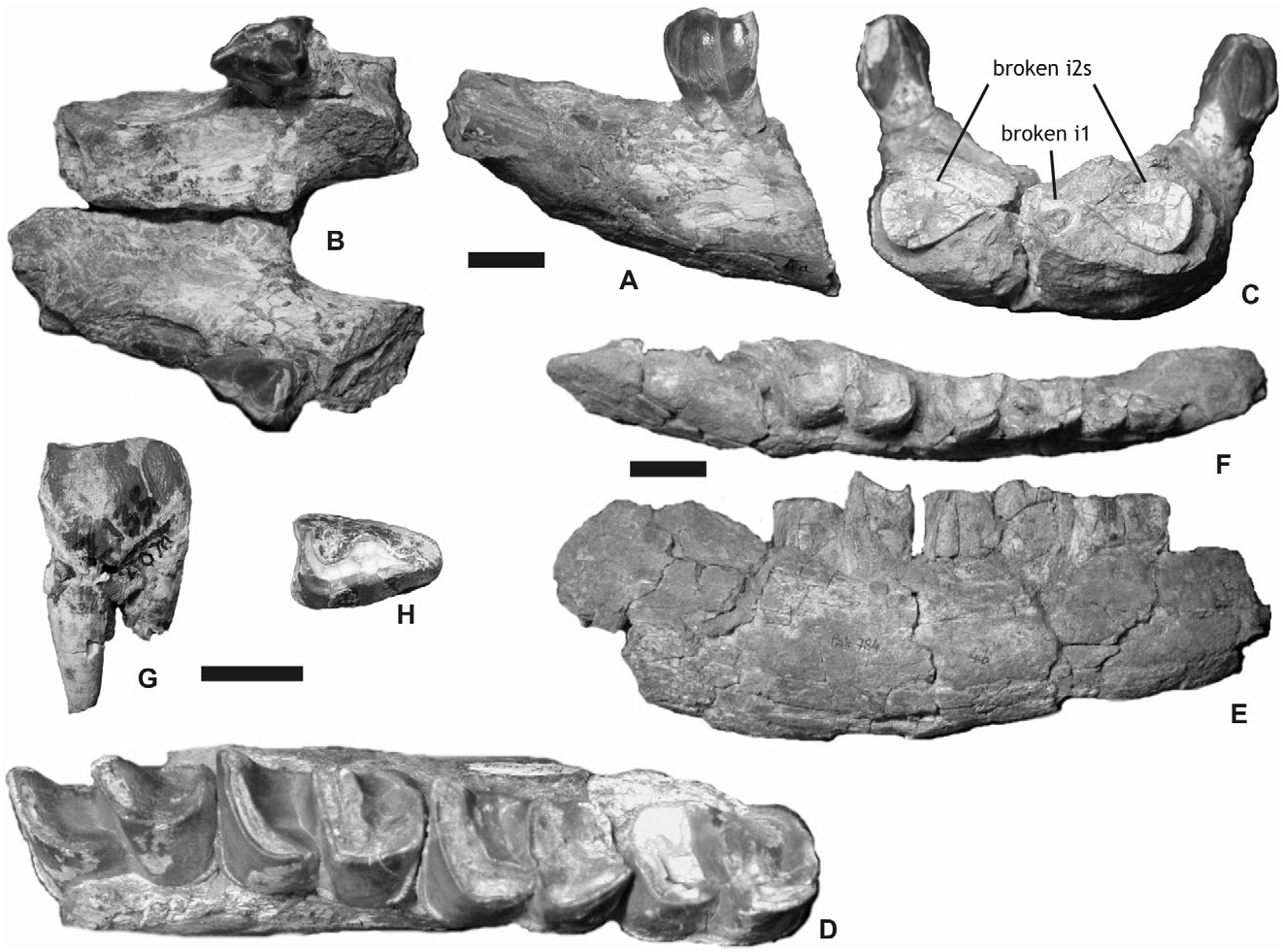
Cranial material: The adult skull MHNT Pak 46 is fragmentary. The occipital, fragments of maxillae, squamosals (processus zygomaticus), and a postglenoid apophysis are preserved ( Fig. 4A–D View Figure 4 ). The palate NHM M 15365 shows identical features. The foramen infraorbitalis was located above P4 and the nasal incisure above the middle of P3. The anterior border of the orbit is above M1. The zygomatic arch was high and thick ( Fig. 4D View Figure 4 ). No processus postorbitalis is present on the dorsal border of the processus zygomaticus of the squamosal. The squamosal–jugal suture is smooth and regular. The caudal border of the processus zygomaticus is depressed by a transverse gutter-like groove ( Fig. 4D View Figure 4 ). The articular tubercle is salient and regularly convex. The postglenoid apophysis, straight in anterior view, has a triangular cross-section, with a convex articular surface. The nuchal tubercle is prominent. The caudal border of the occipital crest is slightly concave. The frontoparietal crests converge rostrally, but their junction cannot be observed on this fragmentary specimen. The dorsal half of the occipital side is depressed. The dorsal side of the skull was narrow: the occipital crest is about 100 mm wide. The junction between the nuchal and temporal crests was very close to the auditory pseudomeatus. The foramen magnum is ·
H, height; L, length, M. w.; M. welcommi ; P. b., P. blanfordi ; post, posterior; proc. coron., processus coronoideus; TD, transverse diameter. Approximate dimensions appear between brackets.
subcircular. A horizontal median ridge splits the occipital condyle into two parts. No medial truncation is visible on the latter.
Mandibular material: The most complete specimen is the mandible MHNT Pak 1038 ( Fig. 5A–D View Figure 5 ). The symphysis, nearly horizontal, forms a plateau continuing the corpus mandibulae. It is thick and rather wide, lacking any lateral constriction at the diastema level. In anterior view, the lingual border of the symphysis is regularly concave and the ventral border is flattened, without a median depression. A sharp and winding ridge separates the lingual and labial borders of the symphysis, between the i2 and the lingual side of p1. The posterior border of the symphysis reaches the middle of p2, as does the wide foramen mentale. The latter is associated with accessory foramina, all along the ventral border of the symphysis. The spatium intermandibulare is very wide – from 32–40 mm between p2 and p3 according to the specimen. In frontal view the tooth rows are strongly divergent, the corpus mandibulae being very oblique ( Fig. 5C View Figure 5 ). The corpus mandibulae gets regularly higher from the symphysis to m1, and notably lower backwards (MHNT Pak 1038, IMC C. 271; Table 1). There is no median sagittal groove (sulcus mylohyoideus) on the lingual side of the corpus mandibulae either in adults (MHNT Pak 1037, 1038, and 1068) or juveniles (MHNT Pak 784, IMC C. 267). The ramus is unknown, but Lydekker (1884: 6, fig. 2) figured a prominent angulus mandibulae (IMC C. 271). The mandible MHNT Pak 784 bears the alveolus of d1, the functional d2–4 series and the m1 included in the pars molaris ( Fig. 5D–E View Figure 5 ). It belongs to a calf, referred to P. blanfordi owing to the shape of m1.
Dental material: The upper incisors are not known with certainty, but the flat wear surface on i2s is most probably because of large I1s. However, the mandibular symphysis MHNT Pak 1038, which is broken in its anterior part, bears cross-sections of the left i1 and both i2s ( Fig. 5C View Figure 5 ). The former has an oval crosssection (5.5 ¥ 7 mm) and it was located below the horizontal line defined by the i2s. The i2s, 22 mm away one from another, have a drop-shaped crosssection, as do the complete i2s MHNT Pak 1021 and 1031. A thin layer of enamel covers the crown. This enamel is fluted in the labial part of the crown. The i2s do not diverge.
The cheek teeth formula is 4P–3M, 4p– 3m. No P1 or persistent D1 can be referred to this taxon. However, the maxilla MHNT Pak 46 bears the broken roots of a small triangular tooth in front of P2 ant, anterior; H, height; L, length; Lect, length of the ectometaloph; P. b., P. blanfordi ; P. p., P. pleuroceros ; post, posterior; W, width.
( Fig. 4B View Figure 4 ). The premolar row is short with respect to the molar one [(LP 3–4/ LM 1–3) ¥ 100 = 46]. There is no enamel folding ( Fig. 4F, K View Figure 4 ). The cement is abundant, covering the ectolophs and filling the valleys. The enamel is thinly wrinkled vertically and even squared because of horizontal striae (MHNT Pak 751, 1024, 1058; NHM M 15337). The crowns are high but still conical, with a strongly oblique ectoloph. The roots are thinly joined, long, and divergent ( Fig. 4A View Figure 4 ).
P2 is trapezoid, wider in its posterior part ( Fig. 4F–G View Figure 4 ; Table 2). The P 3–4s are rectangular, wider than long. The labial cingulum is generally absent (ten P2–4 out of 11), but it forms a low ridge on the P3 MHNT Pak 1024 ( Fig. 4H View Figure 4 ). The lingual cingulum is always strongly developed on P2–4 ( Fig. 4F View Figure 4 ). Generally interrupted on the protocone and/or the hypocone (13/ 16 specimens), it can be continuous (three specimens out of 16). The crochet is lacking on every available P2 ( Fig. 4G View Figure 4 ) and two worn P3s ( MHNT Pak 758, 845). Yet, it is present and always multiple on the eight remaining P3s and the ten available P4s. The crochet is restricted to the top end of the crown, therefore vanishing with wear ( Fig. 4B, F–H View Figure 4 ). The metaloph is complete, V-shaped in occlusal view, and without a constriction. The postfossette is narrow and deep. The median valley is still deeper. The antecrochet, lacking on P2–3, is always strongly developed on P4 ( Fig. 4I View Figure 4 ). There is a lingual bridge joining the lingual cusps on P2–3 and joining the antecrochet with the hypocone on P4 ( Fig. 4F View Figure 4 ). This bridge is thin in Early stages of wear, and it thickens in later stages. On P 2, the metaloph is transverse and the protocone is less developed than the hypocone. The protoloph is narrow but continuous on every P2. There is no medifossette on the upper premolars ( Fig. 4B, F–I View Figure 4 ), except for P4 MHNT Pak 1964. On P 3–4, the anterior constriction of the protocone is generally present; the continuous metaloph forms a dihedron open backwards, in which the crochet is the anterior angle; the hypocone is posterior to the metacone. On P 3, the protoloph is continuous and there is no crista. The parastyle is sagittal. The paracone and the metacone folds are always present on P2–4, the former being thicker .
The upper molars are generally lacking a labial cingulum: only two M2 have a cingular bulge very reduced, at the neck. The antecrochet and the crochet are always well developed, except on worn teeth, where the crochet may vanish (MHNT Pak 1031). The anterior constriction of the protocone is always deep, both in M1–2 and M3 ( Fig. 4E–F, K–M View Figure 4 ). Therefore, the protoloph is ‘trefoil-shaped’ (sensu Antoine, 2003). The crochet is sagittally orientated and generally simple on the upper molars (11 specimens out of 15). Yet, the crochet is sometimes double in the top of the crown (MHNT Pak 760, 1012a, 1045, 1258). No crista ant, anterior; H, height; L, length; P. b., P. blanfordi ; P. p., P. pleuroceros ; post, posterior; W, width. Approximate dimensions appear between brackets.
is present on M1–3, except on M2 MHNT Pak 1027. There is neither medifossette nor cristella. The lingual cingulum is generally reduced, determining a tubercle more or less developed, located at the entrance of the median valley. It forms a thin ridge on the protocone of two M3 (MHNT Pak 761, 918), but is absent from other molars (MHNT Pak 1019, 1022). The ectoloph is nearly straight on M1–2, with a sagittal parastyle, a weakly developed paracone fold, a weak mesostyle but no metacone fold. The metaloph is long on M1–2. A deep constriction notches the anterior side of the hypocone. This constriction is restricted to the base of the crown, deeper on M1 than on M2 ( Fig. 4K–L View Figure 4 ). It is absent on M3, except on MHNT Pak 918. There is a shallow groove on the posterior side of the hypocone, close to the lingual tip of the posterior cingulum on M1–2. There is no junction between the antecrochet and the hypocone, even on worn molars. The postfossette is always present, deep and narrow. No lingual groove notches the protocone of M2. On M3, the ectoloph and the metaloph are fused into an ectometaloph ( Fig. 4M View Figure 4 ). The posterior cingulum forms a thick spur restricted to the lingual half of the latter. Yet, M3 has a trapezoid outline, with a wide posterior side – corresponding to the remnant metaloph – supported by two divergent roots. The protoloph of M3 is sagittal and transverse.
The morphology of p1 is unknown. However, the size comparison between the alveolus of d1 (juvenile mandible MHNT Pak 784) and the one present in the adult mandible MHNT Pak 1038 leads us to assume the occurrence of true p1s in adults ( Table 3). This p1 was single-rooted, with a cylindrical root section. The ectolophid of p2 is covered by vertical rugosities continuing the labial cingulum (PMNH Z2070, Fig. 5A, G View Figure 5 ). On p3–4, such rugosities are replaced by a sinuous and continuous cingulum. The external groove is shallow, U-shaped on every lower cheek tooth, vanishing above the neck. The trigonid is angular and forms a right or obtuse dihedron ( Fig. 5D View Figure 5 ). The metaconid is constricted, contrary to the entoconid. The posterior valley is wide and V-shaped. The lingual cingulum, always present, is restricted to the anterior part of the lower cheek teeth. Continuing the anterior cingulum, it forms a thick ridge interrupted at the metaconide level. The labial cingulum is high and continuous on lower premolars, and reduced on lower molars, forming a short ridge in the external groove. The p2 has an isolated spur-like paralophid ( Fig. 5B View Figure 5 ). The paraconid is developed and globular. The posterior valley of p2 is open. The base of the metaconid – between the roots – is depressed on the available specimens. The hypolophid of the lower molars is oblique. There is no lingual groove on the entoconid of m2–3.
The juvenile mandible MHNT Pak 784 bears no alveolus for deciduous incisors. d1 is one-rooted. The deciduous teeth are damaged, but the metaconid and the entoconid seem to be constricted ( Fig. 5E–F View Figure 5 ). It is impossible to observe the protoconid fold. d2–4 lack both labial and lingual cingula and external roughness. There is no ectolophid fold, but an anterior groove is present on the ectolophid of d2–3. The paralophid of d2 is simple and spur-like. The posterior valley of d2 is lingually open, but a thick oblique ridge lays posteriorly to the metaconid. The paralophid of d3 is double. There is no lingual groove on the entoconid of d3–4.
Postcranial skeleton: The material is very abundant, particularly in the Kumbi 4a locality. Postcranials are small- to medium-sized, very homogeneous in size and proportions ( Tables 5–7, 9–13, 15–21, 23 – 27).
From the humerus, only two distal fragments are referred to this taxon (MHNT Pak 1085, 1198). The fossa olecrani is high. The trochlea is very constricted in its median part. The lateral lip is narrow (TD). Available dimensions are (mm): APD distal extremity = 86; APD trochlea = 71 (medial) (42) (middle), and 49.5 (lateral). The epicondyle is weakly developed and lacking any distal gutter.
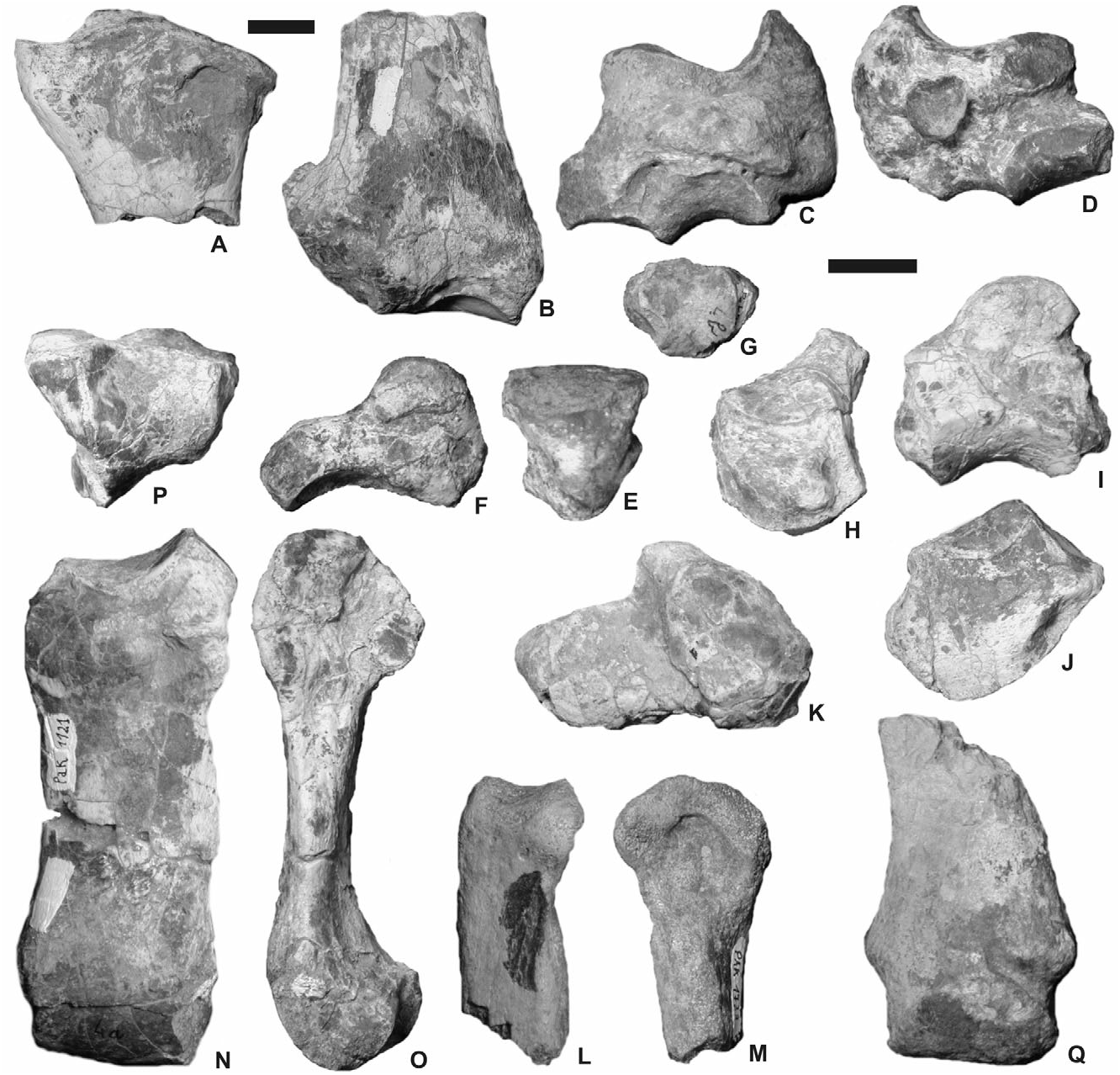
No complete radius is preserved, but proximal and distal fragments are available ( Fig. 6A–B View Figure 6 ; Table 5). In anterior view, the proximal end is much wider than the shaft. The proximal border is sigmoid, with a low medial border and high median part and lateral border. The weak insertion for the m. biceps brachii is slightly depressed medially. In proximal view, the anterior border of the proximal end is straight and the lateral lip of the cochlea forms a deep basin. The proximomedial ulna-facet is low and halfmoonshaped. The proximolateral ulna-facet is high and concave. It is impossible to state whether they are fused or separate. Ulna and radius are independent all along the diaphysis. The distal end of the radius MHNT Pak 1090 ( Fig. 6B View Figure 6 ) bears a huge lateral expansion, which supports the ulnar articular surface and takes the diaphyses away one from another. The diaphysis is slender and dorsoventrally flattened. The gutter for the m. extensor carpi is wide and deepened by a strong anterolateral tuberosity, above the semilunate-facet. The distal end is wide (TD) and flattened dorsoventrally. In anterior view, the distal border is oblique, much lower medially than laterally; the limit between the scaphoid- and the semilunatefacet is marked by a salient ridge. The scaphoid-facet, deep and sagittally shortened, is posteriorly extended by a medial high and triangular expansion. On the lateral side of the distal end, only one ulna-facet is present. Based on the available specimens, it is impossible to state the presence/absence of a pyramidal-facet.
The ulna is unknown.
The carpus is massive, with thick tuberosities and muscular insertions, especially on the scaphoid, the magnum and the unciform ( Fig. 6C–K View Figure 6 ). The scaphoid is low and robust, with a large transverse diameter ( Fig. 6C–D View Figure 6 ; Table 6). Its posterior height widely exceeds the anterior height. The medial side, short of articular facets, bears a very salient tuberosity in its posterior half. Such a tuberosity extends beyond the trapezium-facet. The latter is small and vertically developed. The scaphoid lacks a posteroproximal semilunate-facet, which is replaced by a thick tubercle. The trapezoid-facet is very wide (TD> APD). The magnum-facet, triangular, is sagittally concave and convex transversally. Its anterior end is located very rostrally with respect to the proximal articulation.
The semilunate MHNT Pak 1101 has no ulna-facet, which indicates the presence of a pyramidal-radius articulation ( Fig. 6E–F View Figure 6 ). The proximal facet is convex and short sagittally. The posterior border of the distal pyramidal-facet is twisted posteriorly. The anterior side is smooth, with a rounded distal border. The magnum-facet does not reach the anterior side.
No pyramidal is preserved, neither the pisiform nor the trapezium.
The trapezoid is small and robust ( Table 9). Its anterior side is as wide as high, with a proximal edge regularly convex ( Fig. 6G View Figure 6 ).
The available magnums are broken: no posterior tuberosity is preserved ( Fig. 6H–I View Figure 6 ). The anterior side is as wide as high ( Table 10). Its proximal border is straight in anterior view. The articular process for the semilunate is semicircular in lateral view (diameter = 20 mm). On the medial side, the articular facets are not well separated, the anterior incisure being very shallow. Both facets form subvertical strips elongated sagittally. On the lateral side, the unciform-facet is rectangular and narrow sagittally. The distal McIII-facet is trapezoid.
The unciform is represented by four specimens, of which three are complete ( Fig. 6J–K View Figure 6 ). This bone is low and wide in anterior view ( Table 11). A strong tuberosity lies along the distal border of the anterior side; this tuberosity is more developed medially. The pyramidal- and McV-facets are always independent but close, especially on MHNT Pak 1113. The posterolateral expansion of the pyramidal-facet is present in this only specimen. In anterior view, the semilunate-facet is concave ( MHNT Pak 1113, 1114) or flat ( MHNT Pak 1094, 1112). In proximal view, the posterior tuberosity is slightly longer than the articular part. The former is wide and low. The distal facet has a wide articular surface for the magnum, McIII, McIV, and McV. The latter is regularly concave sagittally and forms an angle about 60° from the horizontal line, indicating a tridactyl manus (i.e. with a vestigial McV) .
The metacarpals are small and robust, sagittally flattened, with wide diaphyses and salient insertions for the m. extensor carpalis ( Tables 12–13).
McII is only represented by an eroded proximal end (MHNT Pak 1733; Fig. 6L–M View Figure 6 ) and a distal half (MHNT Pak 789). The proximal articulation has a quarter-circle outline in proximal view. The magnumfacet is curved in proximal and lateral views. It is impossible to state the presence/absence of a trapezium-facet. The distal end is wide, with a strong lateral tubercle close to the anteroproximal border of the distal articulation. According to the preserved region of the bone, the diaphysis was curved. The distal articulation is very wide, almost symmetrical, and slightly twisted posteromedially. The keel (or intermediate relief) is high and sharp.
Six McIII are preserved ( Fig. 6N–Q View Figure 6 ). One is complete ( Table 13). The proximal end is not widened with respect to the diaphysis. The wide McII-facet is comma-like, elongated sagittally. The magnum-facet is narrow and triangular in proximal view. It is visible in anterior view. The unciform-facet forms a narrow and elongated rectangle triangle. The insertion for the m. extensor carpalis is strikingly salient, with two huge tuberosities (medial and lateral). Thus, the lateral border of the diaphysis is laterally displaced in its proximal quarter (30–40 mm long). In anterior view, the diaphysis is slightly curved inwards, without distal widening. The intermediate relief, high and acute, is visible in anterior view. It is particularly salient in its posterior half. The anteroproximal border of the distal articulation is hardly separate from the diaphysis. No posterodistal tubercle is present on the diaphysis.
McIV and McV are not preserved. However, the tridactyly of the manus can be assumed owing to the shape of the unciform ( Fig. 6J View Figure 6 ), especially from the orientation of the McV-facet, as argued by Antoine & Welcomme (2000) and Antoine (2002).
The coxal and the femur are unknown.
The patella is wide, i.e. as wide as high ( Fig. 7A; Table 15). The muscular insertions are smooth on the anterior side. The most prominent one corresponds to the m. fascia lata. That for the m. rectus femoris is flat. On the articular side, the medial lip is wide, low, and shallow (MHNT Pak 1131). The proximal border of the articular surface is straight. The distal tip is smooth. The lateral lip is weakly notched transversally.
The tibia is represented by five distal ends ( Table 16). There is no anterior groove on the anterior side ( Fig. 7B). An oblique gutter notches the median part of the posterior side. The gutter corresponding to the m. tibialis posterior is always present and is deep and narrow. It is located on the posterior third of the medial side. The tibia and the fibula are independent, as the absence of any synostosis on the lateral border of the diaphysis indicates. The diaphysis has a dropshaped cross-section (MHNT Pak 1126, 1128). The posterodistal apophysis is high and rounded. In distal view, the distal end has a trapezoid outline. The medial border of the cochlea is narrow and very deep. The lateral lip, much wider than the medial one, is almost flat transversally. The distal fibula-facet is semicircular. The contact area does not exceed 40 mm high.
Only one distal half of a fibula can be referred to this taxon (MHNT Pak 1129). The diaphysis is slender, short of any contact with the tibia. The distal end is robust, with a deep lateral gutter for the m. fibularis ( Fig. 7C–D; Table 17). This vertical gutter is located in the posterior third of the head. The astragalus-facet is flat dorsoventrally, subvertical, and slightly concave sagittally.
Eight astragali are preserved ( Fig. 7E–G; Table 18). They are morphologically and metrically homogeneous, wider than high (TD/H = 1.16) and deep (APD/ H = 0.76). The fibula-facet is subvertical and flat transversally. It is very developed anteroposteriorly. The collum tali is high. The posteroproximal border of the trochlea is nearly straight. The trochlea is very oblique with respect to the distal articulation. The lateral lip is very prominent. The laterodistal expansion of calcaneus-facet 1 (sensu Heissig, 1972) is always present and is high and narrow. This facet is very deep sagittally. Calcaneus-facet 2 is flat and oval, higher than wide. Calcaneus-facet 3 is small (MHNT Pak 1140). Calcaneus-facets 2 and 3 are not connected and are separated by a deep notch. On the distal side, the posterior border of the cuboid-facet bears a strong and abrupt inflection. This facet is wide and short. The medial tubercle is salient, overhanging the medial border of the trochlea by about 15 mm.
The four available calcanei are robust, wide, and low ( Table 19). They lack both fibular and tibial facets. The insertion for the m. fibularis longus is marked, forming a deep notch trimmed by a circular ridge ( Fig. 7H). The tuber calcanei is high, slender, and oblique with respect to the processus calcanei. The latter is short and very thick (TD). The beak (rostrum calcanei) is low. The sustentaculum tali is wide. The cuboid-facet is sagittally flat and very deep transversally.
From the second tarsal row, two naviculars, two cuboids, a mesocuneiform, and two ectocuneiforms are preserved. The navicular is low ( Table 20), with a lozengic outline in vertical view ( Fig. 7I). The insertions for muscles and tendons are developed, especially towards the posteromedial tip, which bears a thick tubercle. The proximal articular surface for the astragalus is laterally displaced and splits into two parts because of a sharp transverse ridge (MHNT Pak 1154).
The cuboid is robust, wide (TD), and short (APD; Table 21). The anterior side, square in anterior view, is oblique with respect to a vertical transverse plane. The proximal facet, oval to lozengic, is split into two equal parts by a sagittal groove: the astragalus-facet is more posterior than the calcaneus-facet ( Fig. 7K). The posterior tuberosity is well developed, with an oblique posterior border ( Fig. 7J). The distal tip of this tuberosity overhangs the distal articulation by a few millimetres. The distal MtIV-facet is triangular (MHNT Pak 1158) or trapezoid (MHNT Pak 1159). In both cases, it is deeper than wide (APD> TD).
The mesocuneiform is wide, forming an isosceles triangle in proximal view ( Fig. 7L; Table 23).
The ectocuneiform is high and narrow and is L-shaped in proximal view ( Table 24). It lacks any posterolateral expansion ( Fig. 7M). The distal border is regularly convex in anterior view.
The metatarsus is more slender than the metacarpus ( Fig. 7N–V). The lateral metatarsals are almost as developed as MtIII. The insertions for the m. interossei are short and restricted to the proximal half of the shaft ( Fig. 7N, S–T). Mt II bears a narrow proximal end, sagittally elongated ( Fig. 7N; Table 25). Its outline forms a quarter oval. The mesocuneiformfacet is triangular (isosceles triangle). An oval posteromedial entocuneiform-facet nearly joins the proximal facet. On the lateral side, the anterior MtIII-facet is well developed, flat, and vertical. The diaphysis has a circular cross-section. The distal end is the most robust part of the bone. The distal articulation is roughly square in distal view ( Fig. 7O). It is wide, nearly flat transversally, with a posteromedial expansion. The intermediate relief is salient, especially with respect to the medial lip of the pulley.
No complete MtIII has been recovered, but this bone was rather slender, according to the available fragmentary specimens ( Fig. 7P–Q; Table 26). In proximal view, the anterior border of the articular facet is regularly convex. This facet is only for the ectocuneiform: there is no cuboid-facet. The proximal border of the anterior side is concave in anterior view. There are two flat and well-developed MtII-facets on the medial side of MtIII MHNT Pak 196. On the lateral side, the MtIV-facets are independent. The posterior facet is distally displaced with respect to the anterior one. The diaphysis widens distally (MHNT Pak 1194, 1195), reaching its maximal width (TD) immediately above the distal articulation: two symmetrical tuberosities considerably widen the diaphysis. The intermediate relief is displaced laterally. The medial lip of the trochlea is thus wider than the lateral one ( Fig. 7R). No posterodistal tubercle is present on the diaphysis.
MtIV is robust, with thick ends and a cylindrical diaphysis ( Fig. 7S–V; Table 27). In proximal view, the proximal end is trapezoid, with a right angle defined by its anterior and medial sides ( Fig. 7U). The angle between the other sides exceeds 90°. The articular side is roughly triangular, even if the posterolateral border is strongly convex. The medial border is notched in its median part. The posterolateral tuberosity forms a strip and is split into two equal parts by a tendinous gutter (MHNT Pak 1166). On the medial side, the articular facets are flat and widely separate. They form an angle of about 150°. The posterior facet is oval and sagittally elongated. Its posterior end reaches the posterior tip of the bone. The diaphysis is curved outwards. The brutal curvature occurs at the distal end of the insertion for the m. interossei, in the proximal half of the shaft ( Fig. 7S). The diaphysis is widened by a medial tubercle (muscular insertion), just above the distal articulation. The latter is wide and deeper (APD) than those of the other metatarsals. It is flat transversally in its medial part and concave in its lateral part. The intermediate relief is low and smooth ( Fig. 7V).
Discussion
In the field, the fossil specimens here referred to P. blanfordi represent thus far the most abundant small to medium-sized rhinocerotid specimens, especially in Level 4 (earliest Miocene, c. 22.5 Myr; Fig. 2). The dimensions of the fragmentary skull MHNT Pak 46 fit with those of the palate NHM M 15365 figured by Forster-Cooper (1934: pl. 67, fig. 34) .
Since its initial discovery through dental and fragmentary craniomandibular remains, this species has been referred to half a dozen distinct genera, which range from Recent times ( Rhinoceros and Dicerorhinus ) back to the Late Miocene ( Aceratherium , Chilotherium , and Teleoceras ), and even to the Late Oligocene ( Aprotodon ). However, to our knowledge, no comparison has been made with coeval rhinocerotid genera, such as the teleoceratine Diaceratherium , the aceratheres (sensu lato) Mesaceratherium and Protaceratherium , and the puzzling pair-horned Pleuroceros , which were abundant around the Oligocene– Miocene transition in Europe ( Antoine et al., 2003a). At first glance, the morphological similarity is striking with the type and only species of Pleuroceros , P. pleuroceros , as illustrated and described by Duvernoy (1854 –1855) and de Bonis (1973), especially for the postcranials – which are referred to this taxon for the very first time in the present work.
Compared with the cranial features observable in both P. blanfordi and P. pleuroceros from France (Gannat, Laugnac, and Paulhiac localities; Duvernoy, 1854 –1855; de Bonis, 1973), the Pakistani material is c. 15% larger. Both taxa share a concave occipital crest in dorsal view and they only differ by the horizontal posterior groove on the processus zygomaticus of the squamosal, which is present in P. blanfordi and absent in P. pleuroceros , and by the nasal incision longer in P. blanfordi . The zygomatic arch is high, the processus postglenoidalis is thin and narrow, the frontoparietal crests converge similarly, and the distal border of the nuchal crest is irregular in both species. The occipital side (shape, orientation) and condyle (sigmoid medial border in occipital view) are identical.
The mandible of P. blanfordi is strongly reminiscent of that of P. pleuroceros : all observable mandibular characters are shared by P. blanfordi and P. pleuroceros (e.g. a nearly horizontal symphysis, with sharp sagittal edges running dorsally along the diastema, and a posterior border at the level of p2), with the exception of the lingual mandibular groove.
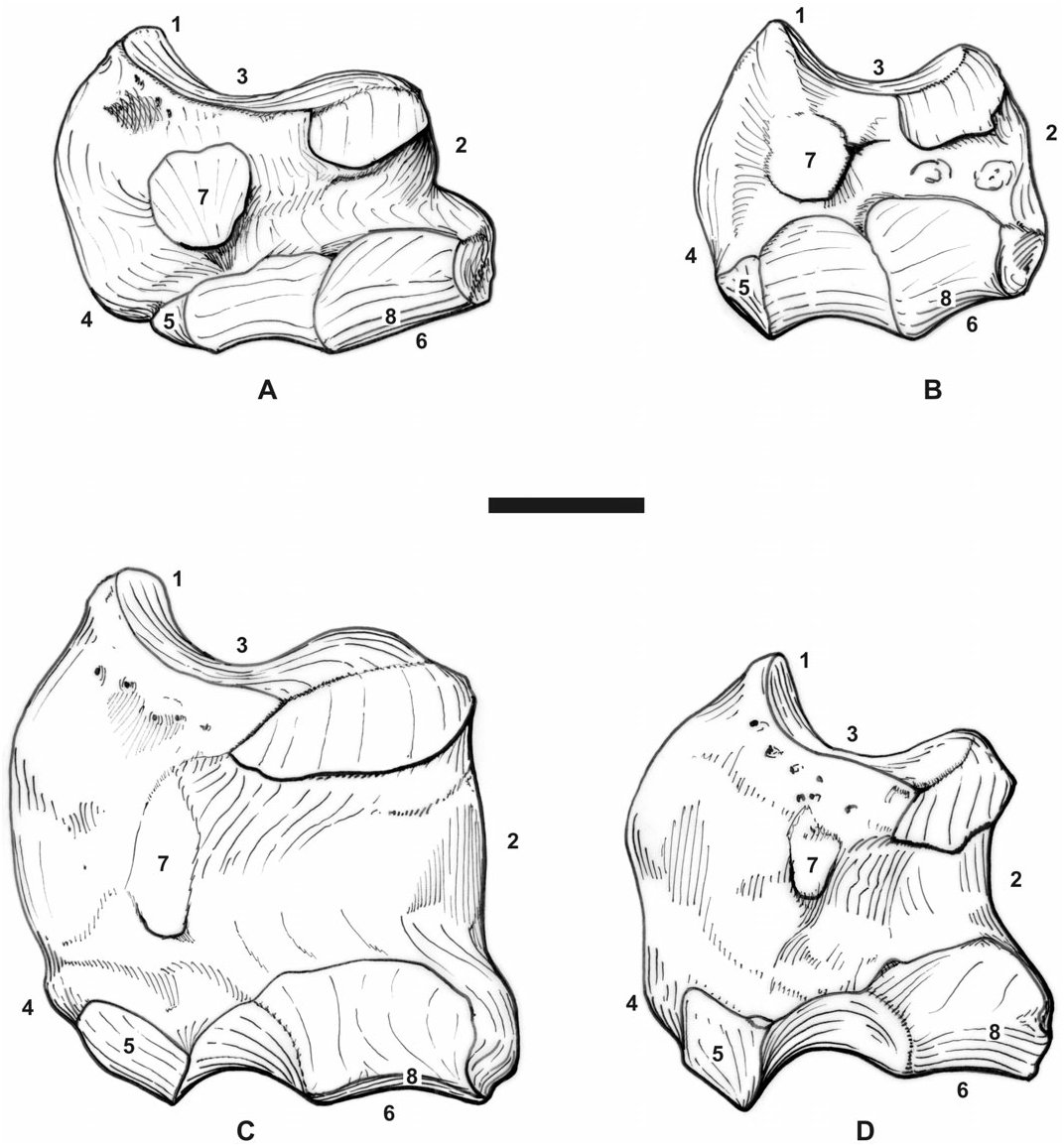
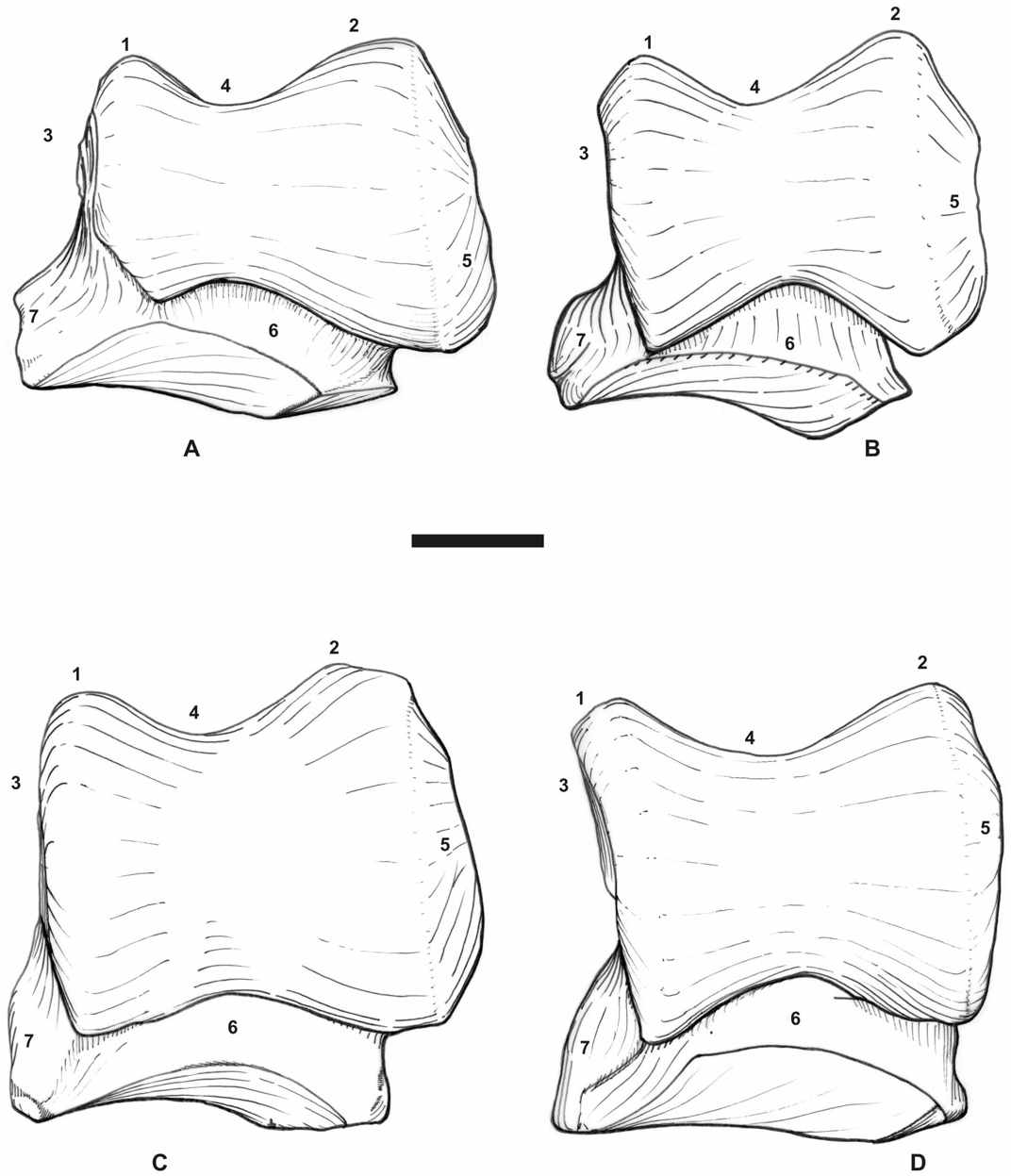
It is still more striking on the postcranial skeleton, with highly similar carpus, tarsus, and metapodials in terms of proportions, articular facets, tuberosities, and trochleae ( Figs 11A–B View Figure 11 , 12A–B View Figure 12 ). It may be noticed that P. blanfordi was most probably tridactyl, as is P. pleuroceros ( de Bonis, 1973: 153, text-fig. 44.7). In both species, the metacarpals have a prominent insertion for the m. extensor carpalis, the tuber calcanei is elevated and slender, and the insertion of the m. interossei on the lateral metapodials is short. The astragali are identical ( Fig. 12A–B View Figure 12 ). The only postcranial differences between both species are the stronger mediodistal tuberosity on the scaphoid in P. blanfordi ( Fig. 11A–B View Figure 11 ) and on McII, the posterior McIII-facet on McII and the fibula-facet on the calcaneus (absent in P. blanfordi ), and the proximal border of MtIII, which is concave in P. blanfordi and straight in P. pleuroceros .
Pleuroceros blanfordi and P. pleuroceros primarily differ in their dental characters, mainly the upper cheek teeth: in P. blanfordi larger dimensions (up to 30% larger; Table 2), a shortened premolar series, higher tooth crowns, abundant coronary cement, a weaker labial cingulum, a multiple crochet (only occasionally observed in P. pleuroceros ), an unconstricted metaloph, a continuous lingual cingulum, the presence of a thick lingual bridge on the upper premolars, a transverse metaloph and a reduced protocone on P2, and the usually constricted protocone on P3–4, the absence of a crista on P3, the unconstricted metaloph on P4, the usual presence of a lingual cingulum (occasional in P. pleuroceros ) on upper molars, a weak paracone fold, and the presence of a metacone fold on M1–2, a strong mesostyle on M2, and a constricted metaconid on the lower deciduous teeth.
Nevertheless, both species share several characters considered as synapomorphies in the phylogenetic analysis performed here (see Phylogenetic relationships): a reduced lingual cingulum on upper premolars, a strong antecrochet on P4, an occasional crista on upper molars, a strongly constricted protocone and a low reduced posterior cingulum on M1–2, a constricted hypocone on M1, a smooth and U-shaped external groove on lower cheek teeth, and a continuous lingual cingulum on lower premolars.
The mandibular symphysis figured by Forster- Cooper (1934: text-fig. 10A) is strongly similar to MHNT Pak 1038, especially for the wide spatium intermandibulare. None of these symphyses is very massive, neither enlarged rostrally nor displaying highly diverging incisors (i2). First lower incisors are retained. The foramen mentale is located under p 2 in P. blanfordi , whereas it is situated in front of it in Aprotodon . An occasional postfossette occurs on the upper cheek teeth. The protoloph joins the ectoloph on P2. The protocone is deeply constricted on the upper molars. The posterior part of the ectoloph is concave on M1–2. A deep anterolingual groove marks the hypocone on M2. The external groove is smooth and U-shaped, and the trigonid is angular and sharp on the lower cheek teeth. The lingual opening of the posterior valley is deep, narrow, and V-shaped on the lower premolars, in lingual view. All of these mandibular and dental features make P. blanfordi differ from the species referred to Aprotodon Forster- Cooper, 1915. To our knowledge, no postcranial remain is referred to the latter genus ( Forster-Cooper, 1915, 1934; Borissiak, 1944; Beliajeva, 1954; Qiu & Xie, 1997).
More features will be discussed in the phylogenetic analysis section, including differences with type species of some genera that P. blanfordi had been referred to, such as Aceratherium and Chilotherium .

| MHNT |
Museum d'Histoire Naturelle Toulouse |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pleuroceros blanfordi
| Antoine, Pierre-Olivier, Downing, Kevin F., Crochet, Jean-Yves, Duranthon, Francis, Flynn, Lawrence J., Marivaux, Laurent, Métais, Grégoire, Rajpar, Abdul Rahim & Roohi, Ghazala 2010 |
Aprotodon
| Metais G & Antoine PO & Baqri SRH & Marivaux L & Welcomme JL 2009: 163 |
Aprotodon blanfordi
| Welcomme JL & Ginsburg L 1997: 1001 |
Chilotherium blandfordi
| Forster-Cooper C 1934: 594 |
Chilotherium blanfordi
| Matthew WD 1929: 508 |
Chilotherium blanfordi Ringström, 1924: 75
| Ringstrom TJ 1924: 75 |
Teleoceras blanfordi Lydekker Pilgrim, 1912: 3
| Pilgrim GE 1912: 3 |
Aceratherium blanfordi var. majus
| Pilgrim GE 1910: 66 |
Aceratherium blanfordi Lyd. Pilgrim, 1908: 149
| Pilgrim GE 1908: 149 |
Rhinoceros blanfordi var. majus
| Lydekker R 1886: 154 |
Aceratherium blanfordi
| Lydekker R 1884: 2 |
Aceratherium blanfordi var. majus
| Lydekker R 1884: 10 |
Rhinoceros palaeindicus
| Lydekker R 1881: 45 |