
Praomys taitae, (HELLER, 1912)
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2012.00817.x |
|
persistent identifier |
https://treatment.plazi.org/id/03B18B5F-2B7D-D10A-FF4B-FACF66841967 |
|
treatment provided by |
Marcus |
|
scientific name |
Praomys taitae |
| status |
|
PRAOMYS TAITAE ( HELLER, 1912) View in CoL ( FIGS 6–9 View Figure 6 View Figure 7 View Figure 8 View Figure 9 ; TABLES 5, 6)
Epimys taitae Heller, 1912: 9 View in CoL ; type locality – ‘ Mt. Mbololo , Taita Mountains [Taita Hills], British East Africa [ Kenya], 5000 feet altitude’ [1524 m]; type specimen – USNM 181797 About USNM , an old adult male collected 5 November 1911 by E. Heller .
Rattus View in CoL [( Praomys View in CoL )] taitae, Hollister, 1919: 79 View in CoL (name combination).
Rattus (Praomys) delectorum, Allen & Loveridge, 1927: 435 (part, provisional identification of series from the Ululguru Mts, Tanzania).
Praomys taitae, G. M. Allen, 1939: 410 View in CoL (name combination).
Praomys jacksoni octomastis Hatt, 1940: 2 ; type locality – ‘ 6000 feet [1829 m] in the Old Mbulu Reserve [Kainam], Tanganyika Territory’ [ Tanzania] ; type specimen – AMNH 55718 About AMNH , an adult female collected 28 November 1928 by A. L. Moses .
Rattus View in CoL [( Praomys View in CoL )] jacksoni View in CoL octomastis, Swynnerton & Hayman, 1951: 316 (name combination, retention as valid subspecies).
Praomys delectorum, Davis, 1965: 131 View in CoL (octomastis and taitae View in CoL allocated as synonyms without indication of rank).
Praomys jacksoni, Misonne, 1974: 27 View in CoL (octomastis and taitae View in CoL allocated as synonyms without indication of rank).
Praomys (Hylomyscus) denniae anselli, Bishop, 1979: 528 (part, misidentification of series from Ngorongoro, Tanzania).
Emended diagnosis: A species of the Praomys delectorum group characterized by small size ( Table 5), delicate molars (CLM ª 4.0–4.2 mm), and relatively heavy incisors; tops of hind feet whitish, lacking dusky swath across metatarsum.
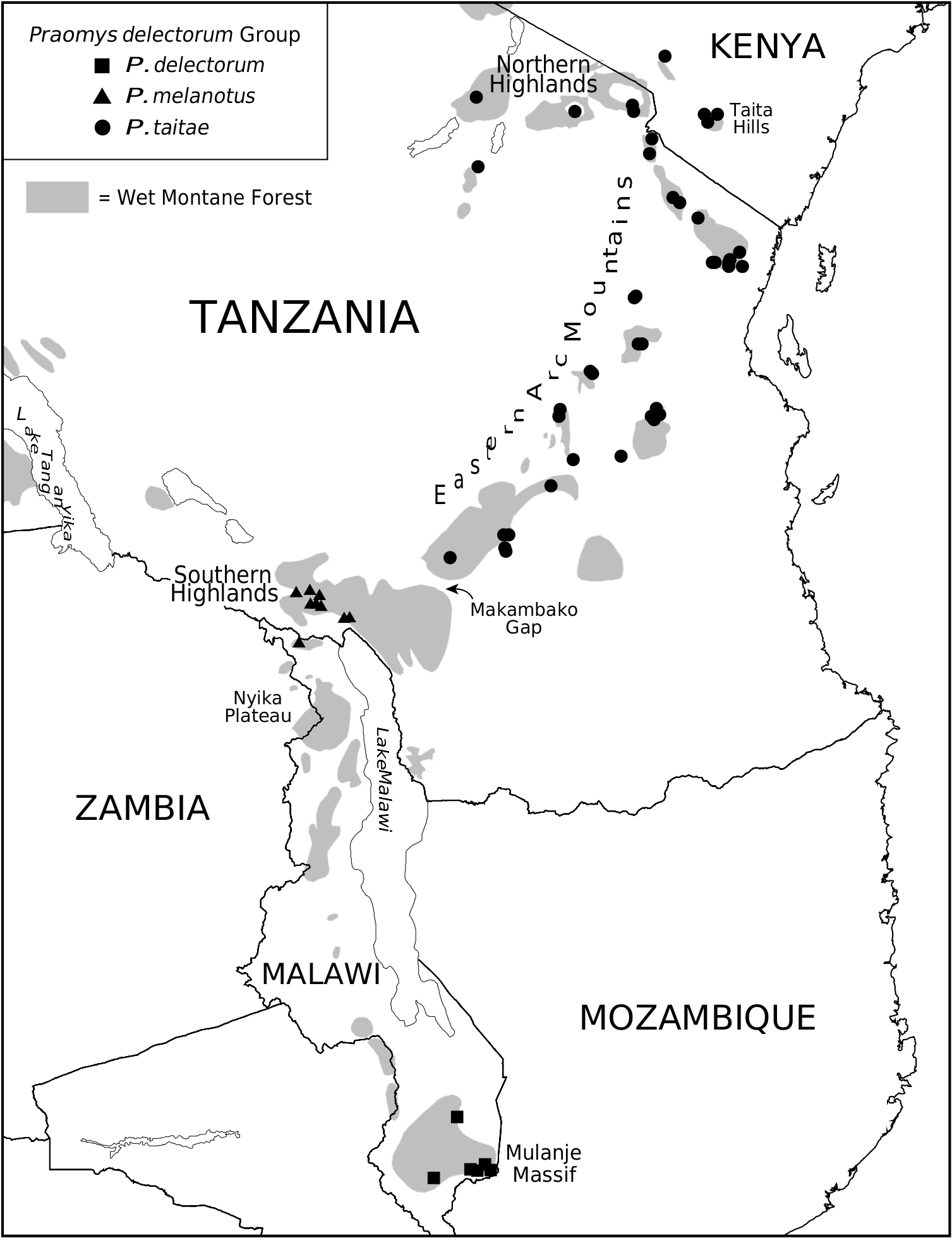
Distribution: Chyulu Hills and Taita Hills of southern Kenya, through the Northern Highlands and Eastern Arc Mountains of northern and central Tanzania, as far south as the western Udzungwa Mts ( Fig. 10 View Figure 10 ); known elevation 230–2895 m, the majority of locality records falling between 1000 and 2000 m.
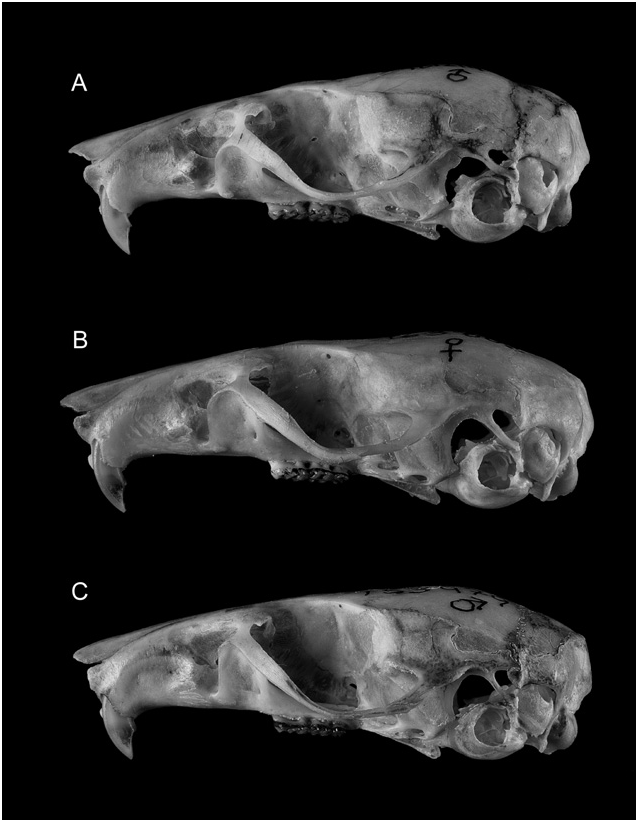
Morphological description and comparisons: Heller’s (1912: 9) characterization of the pelage of P. taitae was brief: ‘Dorsal area russet, darkest medially; sides lighter cinnamon, and sharply contrasted with light underparts; ears and tail broccoli-brown; feet white; underparts whitish, with a cream-buff suffusion; the hairs plumbeous basally’. His chromatic synopsis captures the reddish-brown colours and brighter tone that dominate the pelage in samples of P. taitae , especially compared with those of P. melanotus ( Figs 8A–C View Figure 8 , 9 View Figure 9 ). Some shade of reddish brown covers the crown, back, and rump, tending toward fuscous brown or umber over the middle dorsum and grading to cinnamon or fulvous brown on the cheeks and flanks. The junction of the dorsal–ventral pelage is sharply delineated, without any ochraceous lateral line. Relatively long terminal bands of white or cream partially obscure the lead-grey bases of the ventral hairs, imparting a medium grey to greyish white appearance to the underparts; patches of entirely white hairs occur over the throat and chest in some specimens. Series of P. taitae lack the strong overwash of pinkish-buff that commonly embellishes the underparts of P. delectorum ; compared with series of P. melanotus , the long white tips of ventral hairs give a brighter tone to the grey underparts of P. taitae ( Fig. 8A′ View Figure 8 –C′). Tops of hind feet of P. taitae are uniformly white to dirty white, from the ankles to the tips of toes, standing apart visually from the brown dorsal pelage; this trait usefully contasts with the pes of P. delectorum and P. melanotus , both of which commonly possess a swath of dusky hairs that extend over the tarsus onto the metatarsus ( Fig. 9 View Figure 9 ).
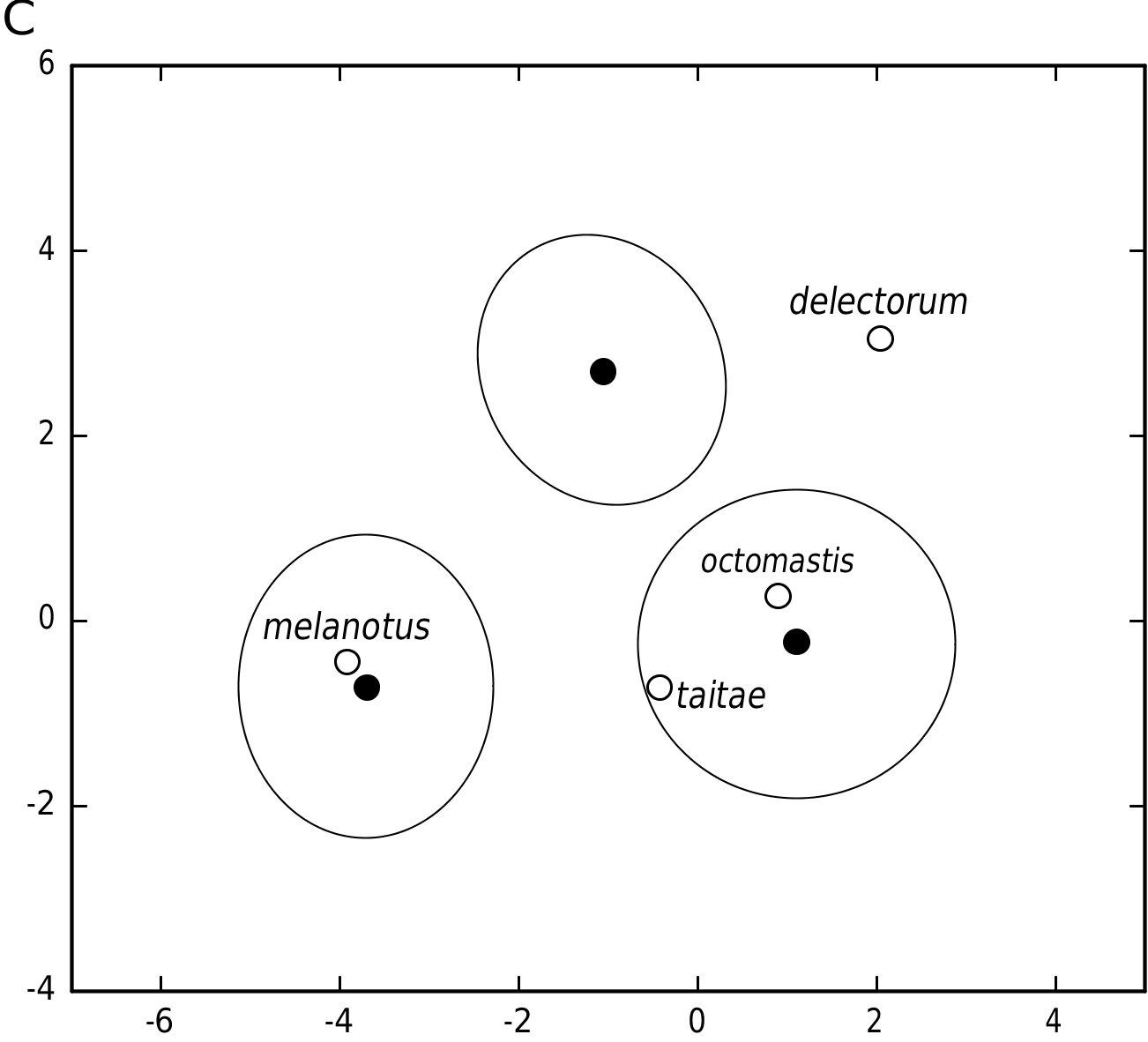
Overall small size is the defining quality of P. taitae among the three species of the P. delectorum group. Its diminuitive size and delicate proportions are attested by the univariate statistics for many variables ( Table 5) and by variable loading coefficients in the multivariate analyses ( Tables 3, 4). Size contrast, unrelated to age, is easily appreciated between samples of the smaller P. taitae and larger P. melanotus ( Fig. 6 View Figure 6 ), but discrimination is less evident between those of P. taitae and P. delectorum . Their cranial differentiation rests upon the narrower zygomatic plate, larger molars, and finely built incisors in P. delectorum compared with relatively broader zygomatic plate, dainty molars, and stout incisors in P. taitae ( Fig. 7 View Figure 7 ).
Remarks: Like Thomas’ (1910) description of Epimys delectorum, Heller (1912: 9) placed his new species, the ‘Taita Forest Mouse,’ within Epimys . Heller (1912: 9) contrasted taitae only with the geographically nearby form peromyscus ( Heller, 1909) – ‘much smaller in size, with shorter ears; skull differs decidedly in lacking beads to interorbital edges, which are rounded, and in the shorter palatal foramina, which reach only anterior edge of first molar’ – the latter currently treated as a synonym of Praomys jacksoni (e.g. Musser & Carleton, 2005). Such trenchant differences served to easily diagnose taitae as distinct from the very different P. jacksoni (peromyscus), but Heller’s description evidenced no awareness of Epimys delectorum Thomas (1910) , then recently described from southern Malawi. Later authors would associate taitae as a syononym of P. delectorum ( Davis, 1965; Musser & Carleton, 1993), and our morphometric evidence indicates its status as a species within the P. delectorum group.
Hatt (1940) considered his new subspecies octomastis to be a distinctive regional form of Praomys jacksoni . At the time (i.e. Allen, 1939), the construct of P. jacksoni was broadly defined to include populations and taxa that subsequent authors have segregated as P. delectorum . As reflected in his choice of the specific epithet octomastis, Hatt (1940: 2) emphasized the mammary formula (2 + 2 = 8) as the ‘only noteworthy point of divergence from surrounding subspecies’. Surrounding subspecies in this case were populations of P. jacksoni peromyscus , which Hatt recognized as possessing only three pairs of mammae (1 + 2 = 6). Hatt (1940: 2) was aware of the description of melanotus by Allen & Loveridge (1933) – octomastis differs from jacksoni melanotus in ‘its generally lighter colour, particularly marked on the under surface’ – but he curiously overlooked their mention (1933: 107) of a nursing female with ‘four pairs of nipples’. All three females of the five specimens forming the type series of octomastis, including the type itself (AMNH 55718), do exhibit eight prominent mammary glands, but we also have verified this count on numerous females within series of P. delectorum , P. melanotus , and P. taitae . The trait is appropriately considered a characteristic of the P. delectorum species group, a contrast that distinguishes it from most members of the P. jacksoni group ( Lecompte et al., 2002a; P. degraaffi also possesses eight teats – Van der Straeten & Kerbis Peterhans, 1999).
Nor did Hatt’s (1940) description of octomastis address Heller’s (1912) taitae , a geographically closer form and one whose morphology essentially resembles octomastis. The type specimen of octomastis (AMNH 55718) agrees closely with that of taitae (USNM 181797) in most craniodental measurements ( Table 6) and nests comfortably within the variation observed for P. taitae , as attested by the mathematical certainty of its a posteriori classification with taitae in DFA ( Fig. 4C View Figure 4 ). The two types exhibit the core qualitative traits of the P. delectorum group – interorbital morphology, supraotic fenestration, long incisive foramina, and pronounced t3 on M1; moreover, the type series of octomastis conforms to the subtle diagnostic traits of P. taitae , notably the wholly white metatarsum, intermediately sized zygomatic plate, smaller molars, and comparatively robust incisors.
As we alluded to above, Allen & Loveridge’s (1933) initial assignment of the Kigogo specimen (MCZ 26498), a juvenile from the Udzungwa Mts, EAM, to melanotus was a dubious allocation. Series from the Udzungwa Mts available to us (see Specimens examined) morphologically fit with other populations in the EAM, the senior name for which is Heller’s (1912) taitae . Although a juvenile, measurements of the unworn molars of MCZ 26498 (CLM = 4.17 mm, WM1 = 1.24 mm) fall at the lowest limits recorded for P. melanotus but within the midrange of values obtained for P. taitae ( Table 5). We regard this past record of melanotus from the EAM as an initial misidentification.
Bishop (1979) allocated specimens from Ngorongoro to the type series of his new subspecies Hylomyscus denniae anselli . Those that we have examined (BMNH 65.3612 –65.3617) prove to be examples of Praomys taitae as indicated by their size, long rostrum and relatively narrow braincase, broad zygomatic plate with deep notch, and well-defined t3 and t9 on the M1.
Although the taxa delectorum , melanotus , and octomastis eventually became associated as geographical representatives of a widely distributed P. jacksoni ( Allen, 1939; Swynnerton & Hayman, 1951; Misonne, 1974), the small Praomys named from the Taita Hills was regularly maintained as a species over this period ( Hollister, 1919; Allen, 1939; Allen & Loveridge, 1942; Swynnerton & Hayman, 1951). Within Tanzania, Swynnerton & Hayman (1951) assigned specimens from the Usambara and Uluguru Mts in the eastern EAM to Rattus taitae , which they maintained as distinct from R. jacksoni melanotus . With reallocation of the populations from the Udzungwa Mts to P. taitae , contra Allen & Loveridge (1933) and Swynnerton & Hayman (1951), the southern distributional occurrence of P. taitae is now well documented, approximately delimited by the Makambako Gap ( Fig. 10 View Figure 10 ). The northern extent of its range is less clear. While the species certainly occurs in the Northern Highlands of Tanzania, we have not found P. taitae in the Gregorian Rift mountains of central Kenya (Mau Escarpment, Aberdare Mts, Mt Kenya), at least in North American collections. Search of other natural history collections is recommended to clarify whether it inhabits those Kenyan highlands.
Natural history notes: Samples of Praomys taitae are generally found in submontane, montane, and upper montane habitats (sensu Lovett, 1993b), but have been documented as low as 230 m in the Kwamgumi Forest Reserve, in lowland forest at the base of the East Usambara Mountains ( Stanley et al., 2005b). The abundance of this species at this elevation (as measured by trap success) was low relative to sites in the same mountain range at higher elevations (Stanley & Goodman, 2011). The Udzungwa Scarp Forest Reserve, Udzungwa Mts, contains undisturbed forest that ranges from 300 to 2000 m, one of the last natural and continuous elevational gradients that can be sampled in the Eastern Arc Mountains. Praomys taitae (reported as P. delectorum ) was recorded from 600 to 2000 m, but in abundance greater than expected at 1460 m; none of the other elevations (600, 910 and 2000 m) yielded numbers of P. taitae greater than expected ( Stanley & Hutterer, 2007). Notably, P. taitae was not found during a survey of the same forest at 300 m of this escarpment a year earlier (W. T. Stanley, unpubl. data). The highest elevation where P. taitae has been documented is 2900 m in forested habitats on Mt Kilimanjaro. Significantly, this species was not observed above the tree line during a faunal survey of this mountain (W. T. Stanley, unpubl. data). Thus, P. taitae is restricted to forests, but ranges lower in elevation than Hylomyscus arcimontensis , with which it is commonly sympatric.
Praomys taitae View in CoL shares its habitat with several other rodent species. Based on intensive faunal surveys over the past two decades, three species have been recorded in every montane setting where P. taitae View in CoL has been recorded: Lophuromys aquilus View in CoL , Grammomys ibeanus View in CoL , and Graphiurus murinus View in CoL . Other species found with P. taitae View in CoL in the Eastern Arc Mountains are Beamys hindei View in CoL and Hylomyscus arcimontensis . The latter two rodents are unknown from the northern highland massifs. The calls for critical systematic evaluation of the various isolated populations of Grammomys ibeanus View in CoL and Graphiurus murinus View in CoL in East African mountains ( Holden, 2005; Musser & Carleton, 2005) advise caution about sympatry among various species until such revisionary studies are conducted.
Data collected over three dry season surveys in the Usambara Mountains indicate that populations of P. taitae View in CoL have limited reproductive activity between July and September. Only 13.5% of females examined (N = 74) were pregnant and only one in 44 (2.3%) females assessed was lactating (Stanley & Goodman, 2011). Between 2004 and 2006, Makundi, Massawe & Mulungu (2006) found a significant density increase in P. taitae View in CoL (reported as P. delectorum ) in the Magamba Forest, West Usambara Mts, in March– June compared with September–December. The greatest number of scrotal testes and perforate vaginas were scored from the period April to June. Based on these data, little reproductive activity occurs during the dry period (July–August) and P. taitae View in CoL apparently breeds annually within a relatively short period. This conclusion is also borne out by data collected during surveys of other Eastern Arc and northern highland populations conducted every year since 1993. In 251 females examined during these collective surveys, only 31 (12.3%) were pregnant. A notable exception is the sample of nine specimens examined in 1996 in the Uluguru Mountains where five (55%) were pregnant. The number of embryos averaged 4.0 (N = 31, range 3–6), the embryos in a single uterine horn ranging from 1 to 6. The embryos averaged 10.5 mm in crown–rump length (range = 2–24 mm, N = 20).
Unlike the scansorial proclivities of Hylomyscus arcimontensis , individuals of Praomys taitae are typically captured in traps set on the ground or in other terrestrial settings, such as on rocks or on logs easily reachable from the ground. Arboreal traps set by Stanley and colleagues commonly caught examples of Hylomyscus arcimontensis , but not those of Praomys taitae . Kingdon (1974) stated that P. jacksoni is partially arboreal.
Specimens examined (365, as follows): KENYA: Nairobi area , Chyulu Hills, 6000 ft ( USNM 344947 About USNM ) ; Coast Region, Taita Hills, Mt Mbololo , Summit , 5000 ft ( FMNH 43458–43460 About FMNH ; MCZ 32124, 32125 About MCZ , 32131 About MCZ ; USNM 181797 About USNM , 183411–183420 About USNM , 183422–183426 About USNM , 183429–183432 About USNM , 184508–184524 About USNM , 184525 About USNM ) ; Taita Hills, Mt Umengo , Summit ( USNM 183433–183441 About USNM ) ; Taita District, Taita Hills, Ngangao Forest , 4.25 km N and 1.75 km W Wundanyi, 1700 m ( CM 102635 View Materials –102643 , 102645–102647 , 102649–102652 ) .
TANZANIA: Arusha Region, Arumeru District, Arusha National Park, Mount Meru , 2300 m ( FMNH 208574–208580 About FMNH , 208583–208585 About FMNH , 208595–208597 About FMNH , 208599 About FMNH , 208601 About FMNH , 208607 About FMNH , 208611 About FMNH , 208612 About FMNH , 208615 About FMNH , 208619 About FMNH ) ; Ngorongoro North , 7500 ft ( BMNH 65.3612 – 65.3617) GoogleMaps ; Old Mbulu Reserve , 5500 ft ( AMNH 55702 About AMNH ), 6000 ft ( AMNH 55715–55718 About AMNH ) . Dodoma Region, Mpwapwa District, Rubeho Mts, Mwofwomero Forest Reserve , 1923 m ( FMNH 197908–197911 About FMNH , 197916 About FMNH , 197918 About FMNH , 197919–197924 About FMNH , 197926 About FMNH , 197927 About FMNH , 197929 About FMNH , 197930 About FMNH , 197934 About FMNH ) ; Mpwapwa District, Rubeho Mts, Mwofwomero Forest , near Chugu Peak, 1900 m ( FMNH 197943 About FMNH , 197945 About FMNH , 197950–197952 About FMNH , 197955 About FMNH ) . Iringa Region, Iringa District, Udzungwa Mts, Ndundulu Forest , 9 km E Udekwa, 1900 m ( FMNH 177709 About FMNH , 177711–177715 About FMNH , 177717 About FMNH , 177719 About FMNH , 177720 About FMNH ) . Kilimanjaro Region, Moshi District, Mt Kilimanjaro, Kilimanjaro National Park , 7 km N and 2.5 km W Maua, 8100 ft ( FMNH 173881 About FMNH , 174269–174271 About FMNH , 174273–174283 About FMNH ) ; Moshi District, Mt Kilimanjaro, Kilimanjaro National Park , 10.5 km N and 3.5 km W Maua, 9500 ft ( FMNH 174285 About FMNH , 174286 About FMNH ) ; Moshi District, Mt Kilimanjaro, 4 km N and 1.5 km W Maua, 6700 ft ( FMNH 174287– 174294 About FMNH , 174296 About FMNH , 174297 About FMNH , 174299–174309 About FMNH ) ; Mwanga District, North Pare Mts, Kindoroko Forest Reserve , 1688 m ( FMNH 192692 About FMNH , 192696 About FMNH , 192697 About FMNH , 192701 About FMNH , 192704 About FMNH , 192705 About FMNH , 192709 About FMNH , 192711 About FMNH , 192719 About FMNH , 192721 About FMNH , 192722 About FMNH , 192728 About FMNH , 192730 About FMNH , 192733 About FMNH , 192734 About FMNH ) ; Mwanga District, North Pare Mts, Minja Forest Reserve , 1572 m ( FMNH 192737 About FMNH , 192738 About FMNH , 192741 About FMNH , 192743 About FMNH , 192744 About FMNH , 192748 About FMNH , 192749 About FMNH , 192750 About FMNH , 192752 About FMNH , 192755 About FMNH , 192762–192764 About FMNH , 192769 About FMNH , 192773 About FMNH ) ; Same District, South Pare Mts, Chome Forest Reserve , 7 km (by air) S Bombo, 1100 m ( FMNH 151311 About FMNH , 151312 About FMNH , 151320 About FMNH , 151335 About FMNH , 151336 About FMNH , 151338 About FMNH , 151339 About FMNH , 151344 About FMNH , 151348 About FMNH , 151349 About FMNH , 151351 About FMNH , 151360 About FMNH ) ; Same District ( FMNH 153972–153974 About FMNH , 153976–153987 About FMNH , 153993 About FMNH , 153996 About FMNH ) . Morogoro Region, Udzungwa Mts , Kigoro ( MCZ 26498) ; Kilombero District, Udzungwa Mts , 3.5 km W and 1.7 km N Chita, along Chita-Ihimbo trail, 910 m ( FMNH 155466 About FMNH , 155467 About FMNH , 155625 About FMNH , 155627 About FMNH , 155630 About FMNH ) ; Kilombero District, Udzungwa Mts , 4.5 km W Chita, along Chita-Ihimbo trail, 600 m ( FMNH 155478 About FMNH , 155653 About FMNH ) ; Kilombero District, Udzungwa Mts , 19.5 km N and 0.5 km W Chita, 2000 m ( FMNH 155480 About FMNH , 155654 About FMNH , 155658–155660 About FMNH ) ; Kilombero District, Udzungwa Mts , 4 km W and 5 km N Chita, along Chita-Ihimbo trail, 1460 m ( FMNH 155631 About FMNH , 155637– 155647 About FMNH , 155649 About FMNH , 155651 About FMNH , 155652 About FMNH ) ; Kilosa District, Malundwe Mts, Mikumi National Forest , 985 m ( FMNH 187296–187301 About FMNH , 187303–187311 About FMNH , 187313– 187318 About FMNH ) ; Kilosa District, Rubeho Mts, Ilole Forest , 1878 m ( FMNH 197956 About FMNH , 197957 About FMNH , 197960–197964 About FMNH ) ; Kilosa District, Ukaguru Mts, Mamiwa-Kisara Forest, 1.5 km S Mt Munyera , 1840 m ( FMNH 166687 About FMNH , 166688 About FMNH , 166944 About FMNH , 166946–166949 About FMNH , 166951 About FMNH , 166952 About FMNH , 166954 About FMNH , 166956 About FMNH , 166957 About FMNH , 166959 About FMNH ) ; Kilosa District, Ukaguru Mts, Mamiwa-Kisara Forest, 1 km E and 0.75 km S Mt Munyera , 1900 m ( FMNH 166685 About FMNH , 166960–166962 About FMNH , 166965 About FMNH , 166966 About FMNH , 166968 About FMNH ) ; Morogoro District, Nguru Mts, Manyangu Forest Reserve , 8 km N and 3 km W Mhonda, 1000 m ( FMNH 161281 About FMNH , 161282 About FMNH , 161284–161286 About FMNH , 161289 About FMNH , 161291 About FMNH , 161294– 161297 About FMNH ) ; Morogoro District, Nguru Mts, Nguru South Forest Reserve , 6 km N and 6 km W Mhonda, 1500 m ( FMNH 161301–161305 About FMNH , 161307 About FMNH , 161310 About FMNH ) ; Morogoro District, Uluguru Mts, Uluguru North Forest , 3 km W and 1.3 km N Tegetero, 1345 m ( FMNH 158366 About FMNH , 158368 About FMNH , 158370 About FMNH , 158371 About FMNH , 158373–158375 About FMNH ) ; Morogoro District, Uluguru Mts, Uluguru North Forest , 5.1 km W and 2.3 km N Tegetero, 1535 m ( FMNH 158379– 158381 About FMNH , 158384–158386 About FMNH , 158388 About FMNH , 158563 About FMNH , 158574 About FMNH ) ; Uluguru Mts, Bagilo ( MCZ 22504, 22506 About MCZ ) ; Uluguru Mts, Vituri ( MCZ 22509, 22512 About MCZ , 22513 About MCZ ) . Tanga Region, Hadeni District, Nguu Mts, Nguru North Forest Reserve , 5.6 km S and 3 km E Gombero, 1180 m ( FMNH 168350 About FMNH , 168354 About FMNH , 168355 About FMNH , 168357 About FMNH , 168358 About FMNH , 168365 About FMNH , 168366 About FMNH , 168368 About FMNH , 168370 About FMNH , 168373 About FMNH , 168374 About FMNH , 168376 About FMNH , 168377 About FMNH , 168379 About FMNH , 168383 About FMNH , 168384 About FMNH , 168386 About FMNH , 163387 About FMNH , 168393 About FMNH , 168394 About FMNH , 168396 About FMNH , 168399– 168401 About FMNH ) ; Hadeni District, Nguu Mts, Nguru North Forest Reserve, 3.6 km S and 4.7 km E Gombero , 1430 m ( FMNH 168402 About FMNH , 168406 About FMNH , 168408 About FMNH , 168412 About FMNH , 168413 About FMNH , 168416 About FMNH ) ; Korogwe Distrtict, West Usambara Mts, 12.5 km NW Korogwe, Ambangulu Tea Estate , 1300 m ( FMNH 147309 About FMNH , 147311 About FMNH , 147313 About FMNH , 147317– 147320 About FMNH , 147327 About FMNH , 147332 About FMNH , 150158 About FMNH , 150280 About FMNH , 150281 About FMNH , 150283–150290 About FMNH , 150352–150354 About FMNH , 151272 About FMNH , 151275 About FMNH , 151513 About FMNH , 151514 About FMNH ) ; West Usambara Mts, 11 km NW Korogwe, Ambangulu Tea Estate , 1120 m ( FMNH 147335 About FMNH ) ; West Usambara Mts, Sunga , 35 mi N Lushoto at Resthouse ( USNM 340787 About USNM ) ; Muheza District, East Usambara Mts, 6 km NW Amani, Monga Tea Estate , 1100 m ( FMNH 150217 About FMNH , 150218 About FMNH , 150221 About FMNH , 150294 About FMNH , 150297 About FMNH , 150307 About FMNH , 150311 About FMNH ) ; Muheza District, East Usambara Mts, 4.5 km WNW Amani, Monga Tea Estate , control site, 1100 m ( FMNH 150236 About FMNH , 150240 About FMNH , 150245 About FMNH , 150250 About FMNH , 150319 About FMNH , 150324 About FMNH , 151280 About FMNH , 151298 About FMNH , 151300 About FMNH ) ; Muheza District, East Usambara Mts, 8 km WNW Amani, Bulwa Tea Estate , 1000 m ( FMNH 150257 About FMNH , 150332 About FMNH , 150337 About FMNH ) ; Muheza District, East Usambara Mts, 4.5 km ESE Amani, Monga Tea Estate , control site, 900 m ( FMNH 150262 About FMNH , 150268 About FMNH , 150274 About FMNH , 150279 About FMNH , 150339 About FMNH , 150346 About FMNH , 150349 About FMNH , 151305– 151307 About FMNH ) ; Muheza District, Kwamgumi Forest Reserve , 4.4 km W Mt Mhinduro, 2 km S Kwamtili Estate offices, 230 m ( FMNH 153998 About FMNH ) ; Magrotto Mt , Magrotto Estate ( MCZ 39057) .



| MCZ |
Museum of Comparative Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Praomys taitae
| Carleton, Michael D. & Stanley, William T. 2012 |
Praomys (Hylomyscus) denniae anselli
| Bishop IR 1979: 528 |
Praomys jacksoni, Misonne, 1974: 27
| Misonne X 1974: 27 |
Praomys delectorum, Davis, 1965: 131
| Davis DHS 1965: 131 |
Rattus
| Swynnerton GH & Hayman RW 1951: 316 |
Praomys jacksoni octomastis
| Hatt RT 1940: 2 |
Praomys taitae, G. M. Allen, 1939: 410
| Allen GM 1939: 410 |
Rattus (Praomys) delectorum
| Allen GM & Loveridge A 1927: 435 |
Rattus
| Hollister N 1919: 79 |
Epimys taitae
| Heller E 1912: 9 |