
Rodriguesiophisis spinifera (Butler, 1876) Butler, 1876
|
publication ID |
https://doi.org/ 10.5281/zenodo.196716 |
|
DOI |
https://doi.org/10.5281/zenodo.5612776 |
|
persistent identifier |
https://treatment.plazi.org/id/03B19568-2507-FFCF-DCE5-C31FFA191C8F |
|
treatment provided by |
Plazi |
|
scientific name |
Rodriguesiophisis spinifera (Butler, 1876) |
| status |
comb. nov. |
Rodriguesiophisis spinifera (Butler, 1876) View in CoL n. comb.
( Figs. 75–87 View FIGURES 75 – 86 View FIGURE 87 ; Tab. 4)
Type locality. Indian Ocean, Mascarene archipelago, Rodrigues.
Holotype. Female. (examined) Rodriguez 7, Gulliver [coll.], BMNH.
Neallotype. Male. Rodrigues, Grande Montagne, Alt. 361 m, 19°42’36’’S, 63°27’48’’E, 21.IV.2009, BIOTAS 2009018, S. Hugel, MNHN (MNHN-ENSIF2641).
Non type material examined. All same as neallotype. Males. 23, 13.II.2008, BIOTAS2008064, S. Hugel leg & coll; BIOTAS2008425, S. Hugel leg & coll. 13, 21.IV.2009, BIOTAS 2009017, S. Hugel leg & coll.. Females. 1Ƥ, 21.IV.2009, BIOTAS 2009019, S. Hugel leg & coll.. 1Ƥ, 26.IV.2009, BIOTAS 2009066, S. Hugel leg & coll..
Redescription. In addition to generic characters.
Pronotum anterior and posterior margins inconspicuously concave. Fore tibia with very small inner and outer ventral apical spurs; with 7/7 subapical spurs. Fore femora with 6 / 5 ventral spurs. Mid tibia with very small inner and outer ventral apical spurs; with 6 / 6 spurs. Mid femora with 4–5 / 1 ventral spurs. Hind tibia with 20–27 anterior dorsal spines; with 18–21 posterior dorsal spines; with 10–12 anterior ventral spines; with 7–8 posterior ventral spines. Hind femora with 14–20 ventral anterior spines; with 3–8 ventral posterior spines.
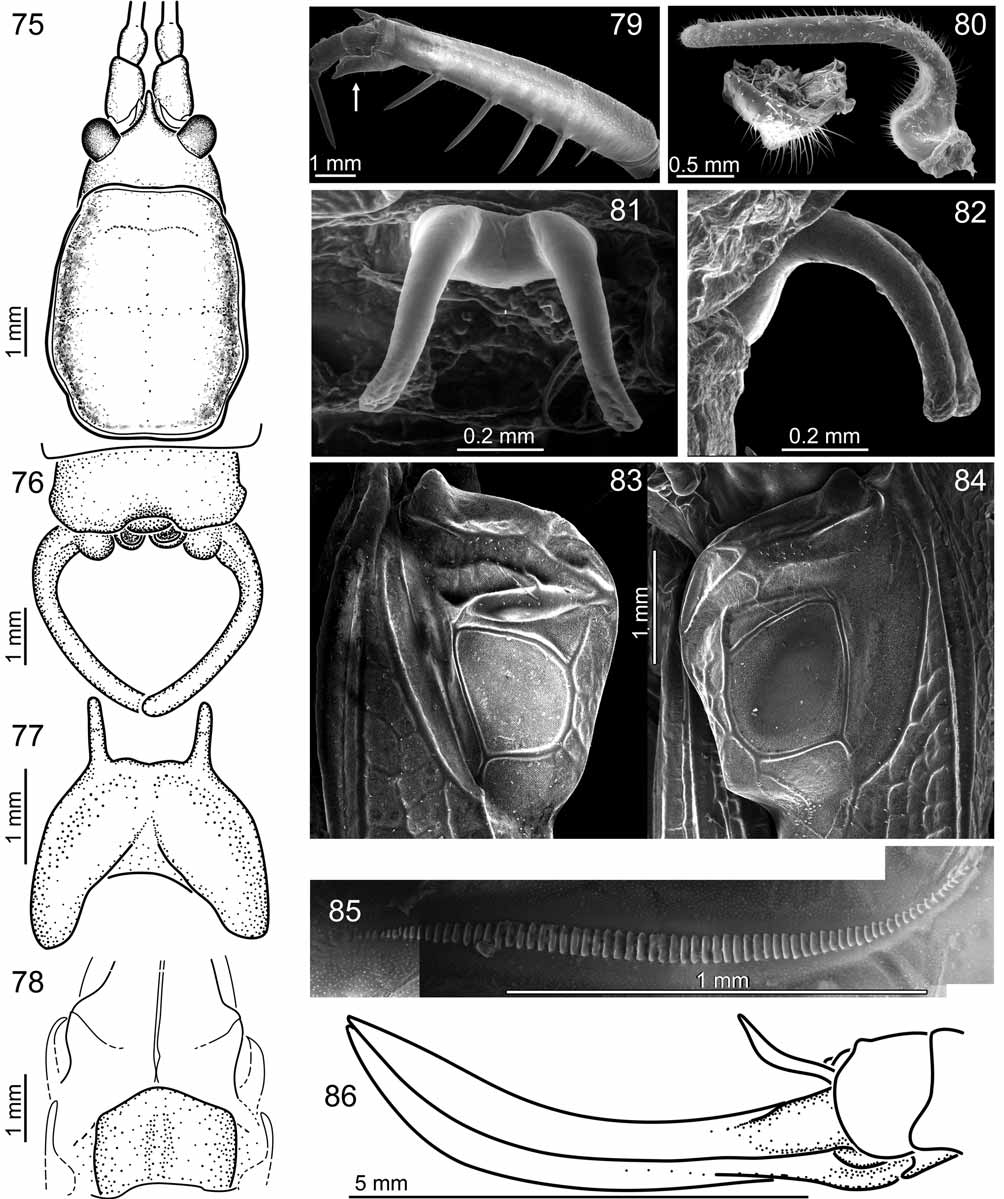
Male. Wings. File with 65–70 lamellar teeth (average: 68, Fig. 85 View FIGURES 75 – 86 ). Terminalia. Epiproct very small, fitting in a notch of the last tergum ( Fig. 76 View FIGURES 75 – 86 ). Paraprocts circular, hardly visible in dorsal view ( Figs. 76, 80 View FIGURES 75 – 86 ). Cerci angled at the basis; bent inwards, with a bulge on the basal bent ( Fig. 76, 80 View FIGURES 75 – 86 ). Subgenital plate posterior margin sinuate, almost strait; with one posterior projection on each side; the styli/projection limit weakly distinct ( Fig. 77 View FIGURES 75 – 86 ). Genitalia. Epiphallus rods regularly arched (in side view, Fig. 82 View FIGURES 75 – 86 ); diverging (in dorsal view, Fig. 81 View FIGURES 75 – 86 ).
Female. Subgenital plate with parallel lateral sides; posterior margin convex ( Fig. 78 View FIGURES 75 – 86 ). Ovipositor curved upwards distally; ventral and dorsal valves weakly serrated apically ( Fig. 86 View FIGURES 75 – 86 ).
Color. Green with darkened file vein (males) and ovipositor apex (females).
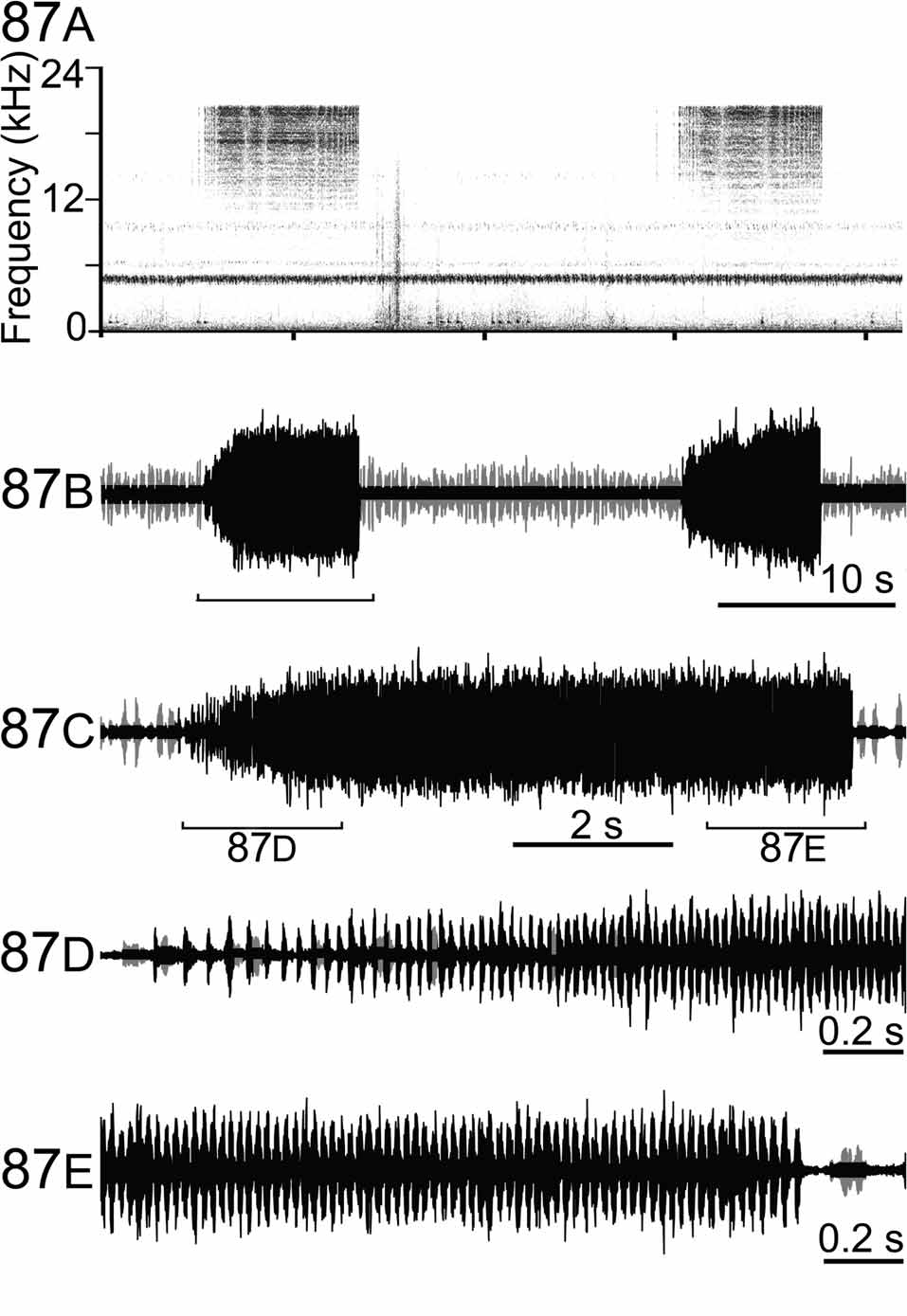
Bioacoustics. Fig. 87 View FIGURE 87 . Males are singing by night hours, often on Latania verschaffeltii . Males are usually moving while calling. Above 20°C, the call of this species consists of short echemes (8.3 ± 0.2 s), with a distinct acceleration of syllables rate at the beginning of the echeme and a deceleration at the very end (unlike the song of the other Mascarene Phisidini species). Syllable rate at the beginning (5 first): 22 ± 2 syllables/s at the middle: 45 ± 1 syllables/s; at the end (5 last): 31 ± 2 syllables/s. Trains of waves forming the syllables are not resolved in natura with our recording setup (3 specimens recorded). The fundamental peaks around 19.5 kHz.
Biology. I directly observed Rodriguesiophisis spinifera in the Grande Montagne nature reserve only, but I recorded one male singing high on an exotic tree ( Syzygium cumini ) in Anse Mourouk nature reserve suggesting that this peculiar species was originally much widely distributed, at least on the humid most part of the island. I observed only juveniles in February and juveniles together with adults in March/April, suggesting that the species might follow a seasonal cycle with adults at the end of the southern summer and during the fall.
In Grande Montagne Nature reserve, the species is rather frequent; it is mostly occurring on the first plots managed by the Mauritius Wildlife Fundation and is more scarce on the plots managed by the Forestry Office (where plants are not confluent yet (in 2009)). Juveniles are staying by day hours on the lowest parts of native shrubs with dense foliage ( Olea lancea , Pittosporum balfourii ) and can be observed by shaking the branches above a sheet or an umbrella; adults seem to spend day hours higher on trees. Most of the singing specimens have been observed walking on the latans Latania verschaffeltii , more rarely on Cassine orientalis . I saw specimens eating small moths and endemic Trigonidiinae yet undescribed (Hugel, in prep.).
Body Head Pronotum Tibia Femora FW O
I II III I II III III
L L W L W H L L L L L L W L L W
3Neallotype 20.6 2.2 3.5 6.2 4.9 2.3 11.2 8.8 16.0 11.5 6.6 14.2 2.0 16.4 ƤHolotype 19.0 9.0 3 (n=4) min 17.1 2.0 3.4 5.6 4.2 2.0 10.2 7.8 16.0 10.4 6.6 13.0 2.0 14.6 max 23.2 2.4 3.8 6.8 5.2 2.3 12.5 9.6 17.0 12.8 8.0 16.5 2.4 17.6 average
19.9 2.2 3.6 6.1 4.7 2.2 11.2 8.7 16.4 11.4 7.1 14.5 2.1 16.0
Ƥ (n=3) min 19.0 2.2 3.1 4.9 3.7 1.6 10.2 8.6 17.1 10.0 7.5 15.2 1.9 17.6 9.0 1.5 max 22.9 2.5 3.5 5.5 4.0 1.9 11.4 9.8 19.3 11.2 7.8 17.3 2.2 20.6 11.9 1.6 average
20.7 2.3 3.3 5.2 3.9 1.8 10.8 9.2 18.2 10.6 7.7 16.2 2.1 19.1 10.5 1.6
Pronotum W: maximal width, including the lateral lobes. Femora W: maximal width. FW W, O W: width on the middle.
| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |