
Saurorhynchus anningae, Maxwell & Stumpf, 2017
|
publication ID |
https://doi.org/ 10.5852/ejt.2017.321 |
|
publication LSID |
lsid:zoobank.org:pub:490DE895-000D-4995-9FAC-1F50DEB0DD0F |
|
DOI |
https://doi.org/10.5281/zenodo.3848070 |
|
persistent identifier |
https://treatment.plazi.org/id/F16C4AFF-4A3B-4C35-8902-F95446F9B1EB |
|
taxon LSID |
lsid:zoobank.org:act:F16C4AFF-4A3B-4C35-8902-F95446F9B1EB |
|
treatment provided by |
Carolina |
|
scientific name |
Saurorhynchus anningae |
| status |
sp. nov. |
Saurorhynchus anningae sp. nov.
urn:lsid:zoobank.org:act:F16C4AFF-4A3B-4C35-8902-F95446F9B1EB
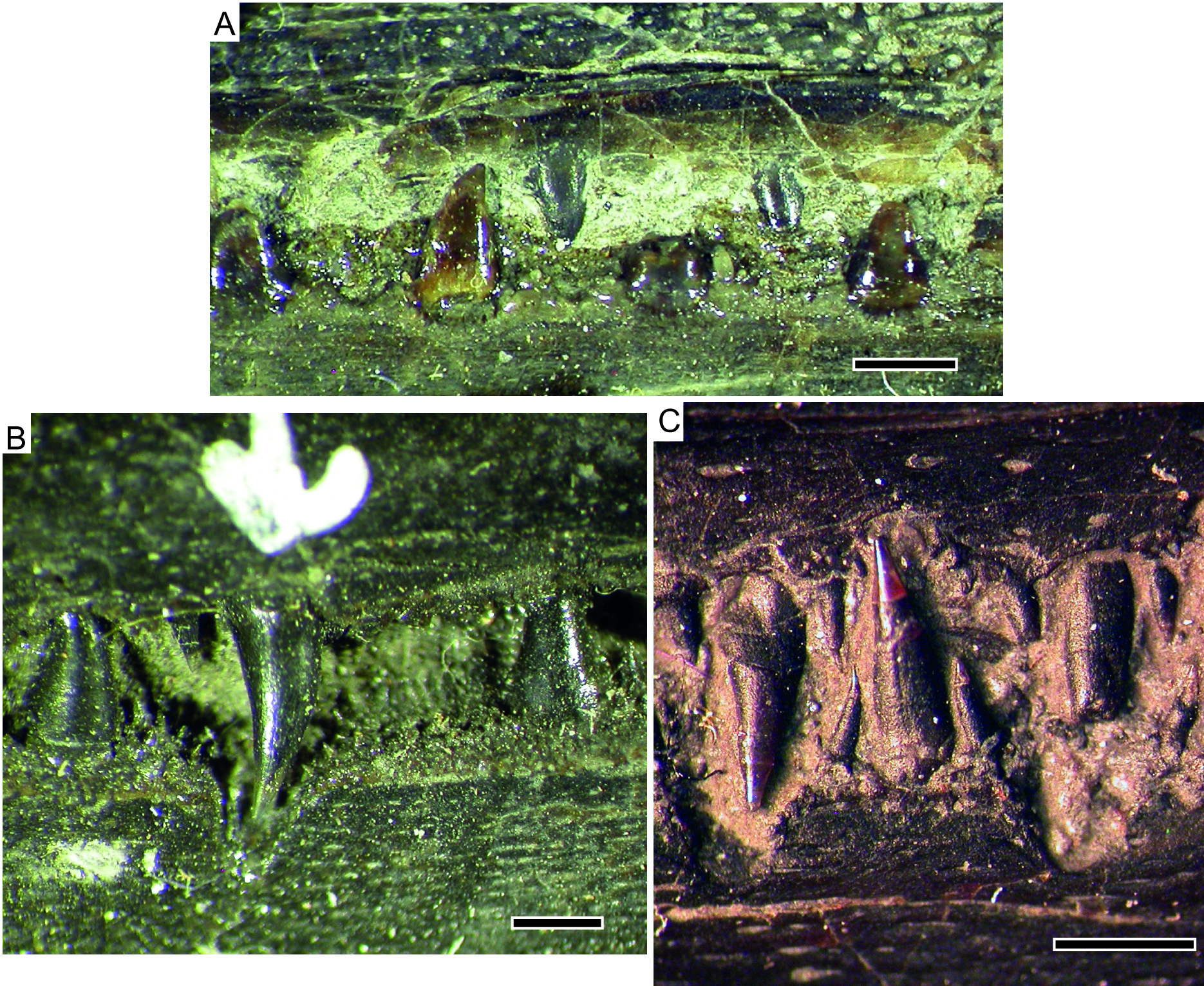
Figs 1C View Fig , 2B View Fig , 3A View Fig , 4A View Fig , 6B View Fig , 7A View Fig
Belonorhynchus acutus – Woodward, 1890: pl. 8, fig. 7. — Woodward 1895, partim: 14–15, pl. 2, fig. 1. Acidorhynchus acutus – Stensiö 1925: 178, fig. 58.
Saurorhynchus acutus – Gardiner 1960, partim: 272–280, figs 19–21. — Forey et al. 2010: 350–352, pl. 65, fig. 5.
Etymology
Name modified from that derived by Agassiz (1844) in honour of Mary Anning, an important fossil collector from Lyme Regis.
Material studied
Holotype
UNITED KINGDOM: a skull and pectoral girdle preserved in lateral view ( NHMUK PV P 3791, Fig. 1C View Fig ).
Stratum typicum
UNITED KINGDOM: Lower Lias. Based on lithological comparisons, most likely the Black Ven Member of the Charmouth Mudstone Formation ( Forey et al. 2010).
Locus typicus
UNITED KINGDOM: Lyme Regis, Dorset.
Differential diagnosis
This species can be differentiated from all other species of Saurorhynchus by the following combination of characters: slender, elongate skull (length to depth greater than in S. brevirostris ); posterior dorsal skull roof essentially parallel to the long axis of the skull; postorbital segment much longer than the depth of the lower jaw (almost double) and double the length of the orbit (unlike in S. acutus ); anterior external narial opening teardrop shaped, widening ventrally; parasphenoid rostrum bearing small denticles ventral to the orbit; foramen for the internal carotid and efferent pseudobranchial arteries displaced posteriorly relative to the basisphenoid and almost entirely enclosed by the parasphenoid (unlike in S. brevirostris , S. acutus ); posterior edge of mandible straight or only weakly sinusoidal, becoming gradually rounded ventrally; angle between the posterior and ventral edges of the mandible greater than 90 degrees; acrodin caps of posterior laniaries not directed lingually (unlike in S. brevirostris ); opercle roughly triangular in shape, tapering ventrally.
Description
The material referred herein to S. anningae sp. nov. has been well-described by Gardiner (1960) as S. acutus . The following description focuses on details in which our interpretations differ from those of Gardiner, or those structures for which additional details can be noted.
The holotype skull is only 89 mm long, however the largest skull referable to S. anningae sp. nov. is 137 mm in length (Suppl. Table 1). Assuming similar proportions to S. hauffi sp. nov., fork length is estimated at 48 cm in the largest individuals.
The antorbital rostrum is composed of the rostropremaxilla, nasaloantorbitals, and frontals. There are two external narial openings, the anterior of which is teardrop shaped and somewhat narrower than reconstructed by Gardiner. The posterior narial opening is small and circular in outline. The suture between the maxilla and premaxilla occurs ventral to the nares. Between the maxilla and the nasaloantorbital is an extremely weakly ossified area that would have borne the infraorbital canal. This space corresponds to the lacrimal, and is more anteriorly extensive than in Gardiner’s reconstruction. Gardiner noted interrostral elements in some of the specimens he examined; we could not confirm the presence of these separate ossifications during restudy of the material. He also noted separate nasal and antorbital ossifications; this observation appears to be based on post-mortem breakage.
The frontal plays a small role in the dorsal edge of the orbit ( Fig. 3A View Fig ). Posterior to the lateral frontal is a small, triangular dermosphenotic, which carries the infraorbital sensory canal and excludes the dermopterotic from the orbit. Ventral to the dermosphenotic is a curved element forming the posterior edge of the orbit and carrying the infraorbital sensory canal; this was identified by Gardiner as the dermosphenotic but is here considered to be the penultimate infraorbital. In some of the most heavily ossified specimens, this element can be quite robust and heavily ornamented.
The lateral extrascapular extends as far anteriorly as the hyomandibula-dermopterotic articulation, unlike in S. acutus , and more anteriorly than in Gardiner’s reconstruction. A dermohyal is present, fused to the lateral surface of the dorsal hyomandibula (as noted by Gardiner 1960). The opercle is also as described by Gardiner.
None of the specimens examined in the present study had a well-preserved ascending process of the parasphenoid, so the posterior position of the foramen for the orbital artery could not be confirmed. Anterior to the ascending process, the parasphenoid bears denticles along its ventral surface. In lateral view, the foramen for the internal carotid and efferent pseudobranchial arteries lies ventral to the opening for the posterior myodome ( Fig. 4A View Fig ), rather than ventral to the anterior basisphenoid as in Toarcian species ( Wenz 1967; Thies 1985).
The mandible consists of three elements in lateral view: the dentary, angular and supraangular. The configuration of the angular-dentary suture reconstructed by Gardiner (1960) is incorrect, as argued
elsewhere ( Griffith 1962). The angular is extensively exposed along the lateral lower jaw, and extends anteriorly approximately as far as the anterior edge of the nasaloantorbital.
The dentary and rostropremaxilla bear large (laniary) teeth, each flanked by a pair of smaller teeth. Laniary teeth are all positioned well anterior to the external narial openings. The posteriormost laniary teeth have relatively straight crowns with acrodin caps, which fit into corresponding pits in the opposite jaw (incisivlücken). Among the posteriormost laniary teeth, the upper teeth overlap the dentary, resulting in incisivlücken on the lateral lower jaw, whereas the lower jaw is slightly narrower and so the laniaries do not project outside the mouth, resulting in an absence of incisivlücken on the lateral surface of the posterior premaxilla ( Fig. 6B View Fig ). The largest teeth are found in the middle of the tooth row; the anteriormost laniary teeth are quite small, similar in size to the flanking teeth. All teeth consist of an acrodin cap, a lightly corrugated region of collar enamel, and an uncorrugated base. Plicidentine appears to be developed around the very base of the tooth, but does not form external ridges above the level of the jawbone. The difference in tooth shape observed between posterior and anterior teeth in S. brevirostris is absent.
Few specimens are available with well-preserved postcrania. The holotype specimen preserves a discrete supracleithrum bearing ornamentation on its external surface ( Fig. 1C View Fig ). A triradiate cleithrum is also present. NHMUK PV P 3790 preserves the most complete postcranium. The neural arch-like elements are small and blocky, similar to those of many saurichthyids. Anteriorly, neural spines are small or absent, and elongate anterior and posterior zygapophyses overlap to form a lattice. In the region around the median fins, neural spines appear to be present, but poor preservation makes this homology interpretation questionable. Haemal spines are preserved in a 1:1 relationship with the neural arch-like elements along the posterior part of the block, dorsal to some poorly preserved anal axonosts. The haemal spines are elongate and bifurcate ventrally. Only a single scale row is present anterior to the median fins, the mid-dorsal scale row ( Fig. 7A View Fig ). The scales are smooth and needle-like, tapering at their anterior and posterior ends. Posterior to the anal axonosts, a mid-ventral scale row is also present.
Remarks
Agassiz (1844: 143 in pt. 2) originally coined the name “ Belonostomus anningiae ” for specimens of Saurorhynchus from the Lower Lias of the UK, but failed to describe the material or identify a holotype. Subsequently, Woodward (1888) figured a non-diagnostic specimen as B. anningiae , but did not provide a description either and subsequently synonymized B. anningiae with Saurorhynchus acutus ( Woodward 1895) . Thus, the name Saurorhynchus anningiae is considered a nomen nudum, and is still available for the longirostrine material from the Charmouth Mudstone Formation (Sinemurian) of the UK as per the original intention of Agassiz.
Occurrence
This species is currently known from the “Lower Lias” of Lyme Regis and Charmouth ( UK) only. Based on lithological comparisons, Forey et al. (2010) surmised that the material of S. anningae sp. nov. at the NHMUK originated from the Black Ven Mudstone Member of the Charmouth Mudstone Formation (? turneri to raricostatum Zones ; latest early Sinemurian to late Sinemurian). The single specimen for which more specific stratigraphic provenance is available, NHMUK PV P 27569 from the Oxynoticeras oxynotum Zone (late Sinemurian), falls within this range.





| NHMUK |
NHMUK |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Saurorhynchus anningae
| Maxwell, Erin E. & Stumpf, Sebastian 2017 |
Belonorhynchus acutus
| Stensio E. 1925: 178 |
| Woodward, 1890 |
| Woodward 1895 |
Saurorhynchus acutus
| Forey P. L. & Longbottom A. & Mulley J. 2010: 350 |
| Gardiner 1960 |