
Saurorhynchus hauffi, Maxwell & Stumpf, 2017
|
publication ID |
https://doi.org/ 10.5852/ejt.2017.321 |
|
publication LSID |
lsid:zoobank.org:pub:490DE895-000D-4995-9FAC-1F50DEB0DD0F |
|
DOI |
https://doi.org/10.5281/zenodo.3848072 |
|
persistent identifier |
https://treatment.plazi.org/id/CF1D94D7-EE54-4E13-B059-97C2536D27FA |
|
taxon LSID |
lsid:zoobank.org:act:CF1D94D7-EE54-4E13-B059-97C2536D27FA |
|
treatment provided by |
Carolina |
|
scientific name |
Saurorhynchus hauffi |
| status |
sp. nov. |
Saurorhynchus hauffi sp. nov.
urn:lsid:zoobank.org:act:CF1D94D7-EE54-4E13-B059-97C2536D27FA
Figs 1 View Fig D–E, 2C, 4C, F, 5C–D, 6C, 7B–D
Belonostomus acutus Quenstedt, 1856 –1858: pl. 29, fig. 8.
Belonorhynchus brevirostris Woodward, 1895: 18 (partim).
Belonorhynchus brevirostris – Woodward 1899: fig. 1
Acidorhynchus brevirostris – Hauff 1938 partim: pl. 22, figs b,c. — Thies (1985) partim: pl. 1, figs 1, 5.
Saurorhynchus brevirostris – Hauff & Hauff 1981: fig. 76. — Urlichs et al. (1994): fig. 79. — Böttcher (1998): fig. 7.15.
Etymology
Named in honour of Bernhard Hauff, whose doctoral dissertation described the saurichthyid fishes from the Holzmaden region.
Material studied
Holotype
GERMANY: Articulated skeleton lacking median and pelvic fins ( SMNS 55057, Fig. 1 D View Fig ). Although the skull of this specimen is not perfectly preserved, it was selected as the holotype based on the presence of extensive postcranial material.
Referred material
GERMANY: all referred material consists of isolated skulls, unless otherwise noted. Bad Boll , εII 10–12 ( SMNS 96878 /1); Dotternhausen , εII 4 ( SMNS 51007 , Fig. 6 C View Fig ); Dotternhausen, εII 6 ( SMNS 58394 ); Haverlahwiese, elegantulum Subzone ( NLMH 70598 ); Holzmaden , εII 3 ( MHH 2 ); Holzmaden, εII 4 ( SMNS 55302 ); Grimmen, exaratum Subzone ( GG 20001, Fig. 3 B View Fig ); Ohmden ( SMNS 53980 , Fig. 5 D View Fig ); Ohmden, εII 3 ( SMNS 50075 ); Ohmden , εII 4 ( SMNS 51888 , Figs 4F View Fig , 5 C View Fig ); Schandelah, exaratum Subzone ( GZG.V.27931); Schandelah, serpentinum Zone , articulated skeleton, caudal region and median fins absent ( NMB 373).
UNITED KINGDOM: Whitby, Toarcian ( NHMUK OR 39153) .
Stratum typicum
GERMANY: Posidonienschiefer Formation, horizon εII4 (uppermost semicelatum Subzone-lowermost exaratum Subzone ).
Locus typicus
GERMANY: Zell unter Aichelberg, Baden-Württemberg.
Differential diagnosis
This species can be differentiated from all other Saurorhynchus species by the following combination of characters: short, deep skull; posterior dorsal skull roof deflected dorsally relative to the long axis of the skull (parallel in S. acutus and S. anningae sp. nov.); postorbital segment approximately equal in length to the depth of the lower jaw (approximately twice as long in S. anningae sp. nov.); overbite slight or absent (unlike in S. brevirostris ); maxilla not ventrally deflected under the orbit (unlike in S. acutus ); coarse, reticular ornamentation on the dermal bones of skull roof (ornamentation reduced in S. acutus ); parasphenoid edentulous ventral to the orbit (unlike in S. anningae sp. nov.); foramen for the internal carotid and efferent pseudobranchial arteries centered ventral to the anterior basisphenoid and oriented laterally (unlike in S. anningae sp. nov. and S. brevirostris ); foramen for the ophthalmica magna artery situated between two ridges on the anterior surface of the basisphenoid (anterior surface of basisphenoid flat and foramen oriented anterolaterally in S. brevirostris ); posterior edge of mandible strongly sinusoidal (straight or only weakly sinusoidal in S. brevirostris and S. anningae sp. nov.); angle between the posterior and ventral edges of the mandible less than 80 degrees (more than 90 degrees in S. anningae sp. nov.); mandibular sensory canal positioned along the dorsal half of the posterior mandible; anterior narial opening ovate (elongate in S. acutus ); subnarial laniary teeth absent (unlike in S. brevirostris ); lateral supracleithrum bearing prominent anterior process pierced by a foramen.
Description
As with S. anningae sp. nov., material now attributed to Saurorhynchus hauffi sp. nov. has been previously described ( Hauff 1938; Thies 1985). However, as much of the material on which the initial descriptions were based is referable to S. acutus , a redescription is necessary. S. hauffi sp. nov. exhibits lower jaw lengths of up to 115 mm (Suppl. Table 1); this would correspond to a fish approximately 40 cm in length.
The rostropremaxilla is the dominant bone in the rostrum, dorsoventrally compressed at the anterior tip and becoming strongly laterally compressed posteriorly. The posterior rostropremaxilla bears a ventral process that forms an overlapping suture with the maxilla ventral to the external narial openings, as well as a dorsal process that approaches the anterior narial opening. Between these two processes, an anterior ventral process of the nasaloantorbital bearing the sensory canal is enclosed. The rostropremaxilla is ornamented with longitudinal grooves. Incisivlücken are lacking at the anteriormost tip. The rostropremaxilla bears approximately 30 laniary teeth. The anterior teeth are very small, and become progressively larger towards the midpoint of the jaw. The sensory pit line runs along the lateral surface of the premaxilla immediately dorsal to tooth row. The sensory pits become increasingly obscured by incisivlücken anteriorly. The presence or absence of interrostrals could not be confirmed.
The maxilla extends anteriorly as far as the first laniary tooth. The suborbital bar is dorsoventrally compressed and bears denticles on its occlusal surface. Posterior to the orbit and anterior to the deepest point of the posterior lamina, a small concave embayment is present along the ventral edge of the maxilla. The postorbital lamina is ornamented with pits, the more posterior of which are elongated in such a way that they appear directed towards the suborbital bar.
The nasaloantorbital is triangular in lateral view. The posterior end encloses two external narial openings: a small, round posterior opening and a large, oval to reniform anterior opening ( Fig. 5C View Fig ). Between the two narial openings runs the supraorbital sensory canal. This canal splits into two branches ventrally; the junction is contained within the nasaloantorbital. Because the lacrimal is often damaged or missing, the ventral edge of the nasaloantorbital is often clearly exposed and has a complex morphology dictated by the orientation of the canals. There is no evidence for separate ossifications of the nasal and antorbital. The nasaloantorbital articulates with the frontal dorsally and the premaxilla ventrally. Ornamentation can be quite strong in the around the narial openings, but becomes reduced anteriorly. Between the posterior narial opening and the orbit is an area of strong dermal ornamentation associated with an element posterior to the nasaloantorbital and excluding it from contact with the anterior orbital margin. This element appears to be anamesic and is tentatively identified as a supraorbital.
The orbital series is heavily ossified. The frontal is not excluded from the dorsal edge of the orbit ( Fig. 3B View Fig ), in contrast to previous reconstructions ( Hauff 1938; Thies 1985). The lacrimal is weakly
ossified and anteroposteriorly elongate, articulating with the maxilla ventrally, premaxilla anteriorly, and nasaloantorbital and supraorbital dorsally. It transmits the infraorbital sensory canal.
The dermosphenotic is variable in size, and forms the posterodorsal edge of the orbit. It carries the infraorbital canal. Ventral to the dermosphenotic, an elongate infraorbital forms the majority of the posterior orbit and carries the sensory canal. Infraorbitals along the ventral edge of the orbit are extremely weakly ossified (just a thin tube of bone surrounding the infraorbital canal, not in direct articulation with more anterior or posterior elements), and as such are rarely preserved.
Posterior to the dermosphenotic and fused with the lateral dermopterotic, an elongate, anamesic element is sometimes preserved. As with the dermosphenotic, it is somewhat variable in terms of size. It is often missing, and even when present the sutural contact can be very difficult to see. This element is most similar to the “supraspiracular plate” of Birgeria groenlandica Stensiö, 1932 in shape, position, and the absence of a sensory canal ( Nielsen 1949). The ventral dermosphenotic in B. stensioei Aldinger, 1931 ( Romano & Brinkmann 2009) was later homologized with the “supraspiracular plate” of B. groenlandica , however the ventral dermosphenotic should contact the infraorbital and carry the infraorbital canal ( Poplin 2004), which the elongate element in S. hauffi sp. nov. does not. We tentatively consider this element to be a neomorph.
The dermopterotic is a broad dorsally convex sheet of bone making up the posterior skull roof ( Fig. 3B View Fig ). A descending process forms an interdigitating suture with the ascending process of the parasphenoid, and creates a notch into which slots the posterior end of the neomorph. Ornamentation generally consists of pits. There is an elongate pore medial to the hyomandibular contact indicating that the medial process of the infraorbital sensory canal was present. Anterior to this canal are the parietals. These form a large, oval ossification. Ornamentation is reticular. No evidence of sensory canals or pores could be detected.
An elongate lateral extrascapular is sutured to the lateral dermopterotic, dorsal to the hyomandibula ( Fig. 5D View Fig ). It transmits the posterior infraorbital canal.A medial extrascapular is also present, and together they form a v-shaped articulation with the dermopterotic.
A dermohyal was present, as the dorsal portion of the hyomandibular bears the pitted ornamentation characteristic of dermal bones ( Fig. 5D View Fig ). A prominent posteriorly oriented joint surface on the ventrolateral dermopterotic articulates with the hyomandibula.
The palate is not exposed in any of the skulls attributable to this species, however the parasphenoid is often well-exposed in lateral view. The parasphenoid rostrum is curved and edentulous ventral to the orbit; the posterior stem is straight and dorsally displaced. The ascending process of the parasphenoid projects dorsolaterally, and is ornamented with fine striations. The foramen for the ophthalmic artery is closer to the anterior than posterior edge of the ventral ascending process ( Fig. 1D View Fig ). There is no expanded tooth plate ventral to the ascending process, and the entire parasphenoid posterior to the anterior margin of the orbit is edentulous. The posterior stem of the parasphenoid is broad. Anteriorly, a central ridge and two lateral ridges are present on the ventral surface of the posterior stem; posteriorly the central ridge diminishes such that it is absent at the posterior end.
Posterior to the orbit, the large foramen for the internal carotid and efferent pseudobranchial artery is contained within the basisphenoid ( Fig. 4C, F View Fig ). In one specimen (SMNS 55302) this foramen is subdivided into anterior and posterior components by a pillar of bone. On the anterior surface of the basisphenoid, the large foramen for the ophthalmica magna artery is recessed in a groove laterally bordered by cristae. Posterior to the basisphenoid, the space between the ascending process and basisphenoid has been interpreted as the posterior myodome. The posterior myodome appears to be roofed by an independent ossification dividing the posterior myodome from the trigeminofacialis chamber. Dorsal to this ossification, the foramen for the ramus oticus lateralis emerges from the neurocranium.
The preopercle is relatively narrow, with the dorsal ramus being narrower than the posterior ramus. It carries the preopercular sensory canal. Unlike previous reconstructions ( Hauff 1938; Gardiner 1960; Thies 1985), it is clear that the preopercular sensory canal runs along the length of the dorsal ramus as well as the posterior ramus, as in Saurichthys . The dorsal edge of the preopercle bears two tiny processes, the anterior of which is directed dorsoanteriorly and the posterior of which is directed anteromedially. These presumably articulate with the hyomandibula. The suture between the posterior ramus of the preopercle and the maxilla is easy to differentiate based on the change in the direction of ornamentation at the suture. The quadratojugal is rectangular in shape and meets the maxilla anteriorly and the preopercle dorsally and posteriorly. It contacts the articular ventromedially.
The lateral lower jaw consists of four elements: a tiny articular, a small supraangular, an expanded angular, and the dentary. The articular is in contact with the supraangular anteriorly. The dorsal portion of the angular is very thin and often eroded, making it appear that the sensory canal is located near the dorsal edge of this element. However, that is not the case. The mandibular sensory canal is contained entirely within the angular, and runs closer to the occlusal jaw margin than the ventral edge. It gives rise to descending canals. The posterior edge of the angular is strongly sinusoidal, and projects posterior to the jaw joint. The angular is also strongly ornamented with pitting and some reticulation; there are a series of “growth lines” running parallel to the ventral edge. The dentary itself has a woody texture, with the grain oriented parallel to the long axis of the mandible. The ventral part of the angular-dentary suture begins as at a posteroventrally angle dorsally, but becomes deflected ventrally.
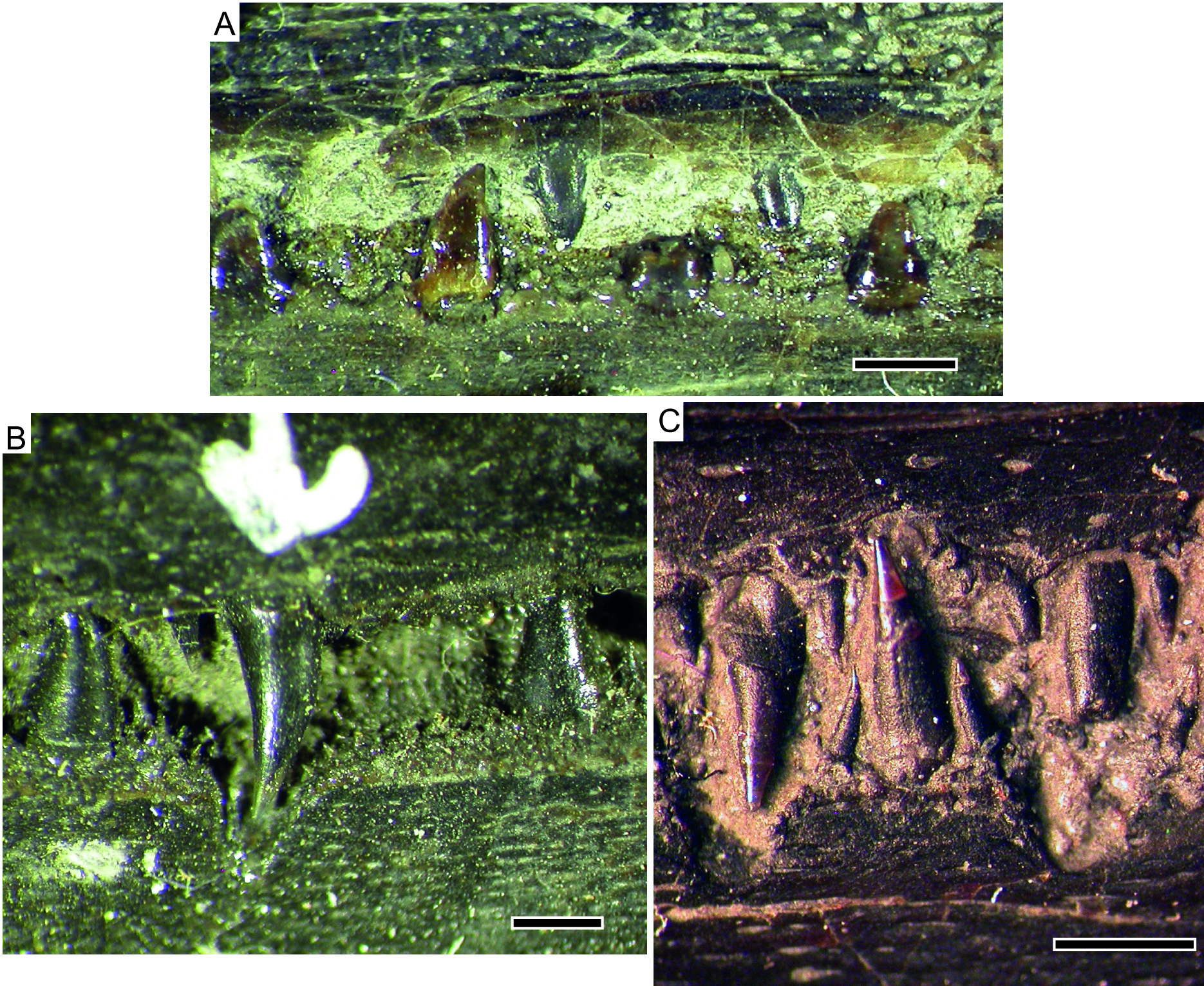
The laniary teeth are simple cones bearing an acutely pointed acrodin cap, and in general appear to be entirely unornamented and lack any differentiation between tooth base and “collar ganoine” ( Fig. 6C View Fig ). However, a single specimen (SMNS 51007) does show extremely faint corrugation of the collar region. We consider that its small maximum tooth size is possibly a reason for restricted expression of corrugation in the collar region (the specimens of S. anningae sp. nov. that clearly show a corrugated collar region are among the largest specimens, far larger than the largest S. hauffi sp. nov.). Some variability in the degree of expression must also be assumed ( Gardiner 1960).
Postcranium
PAIRED FINS. The supracleithra are seldom preserved. They appear to be thin and relatively straight dorsally, ventrally bearing a prominent anterior process that is pierced by a foramen on its medial surface. There is no equivalent to this ventral anterior process of the supracleithrum within Saurichthys , but this morphology is shared with S. brevirostris . The cleithra are triradiate, with a robust posterior ramus. The paired fins are even more rarely preserved. In NMB 373, the pectoral fins are triangular in shape, with the longest and most robust lepidotrichia located close to the leading edge of the fin. A precise count of lepidotrichia is not possible. In no specimens referred to S. hauffi sp. nov. are pelvic fins preserved.
AXIAL SKELETON. The following description is based on SMNS 55057. Sixty-three abdominal and 117 caudal neural arch-like elements are preserved anterior to the caudal fin, for a total of 180 (90 vertebral segments). As indicated by the low abdominal neural arch count, the abdominal-caudal transition is displaced anteriorly relative to the anal fin ( Fig. 1D View Fig ). The neural arch-like elements consist of a short, robust base, a moderately long prezygapophysis and a long, tapering neural spine. The medial surface of alternating neural arches is pierced by a foramen for the intersegmental vessel ( Wu et al. 2015). The neural spines become longer more posteriorly in the column, and become forked immediately anterior to the insertion for the dorsal fin. Posterior to the dorsal fin, the neural spines are not forked for a short distance, but become bifurcated again and posteriorly inclined towards the caudal fin. The haemal spines are short, robust hooks anteriorly, lying dorsal to the gastric mass. Some of them are also bifurcated in this region. They lengthen at the end of the abdominal cavity, and become bifurcated or even trifurcated dorsal to the anal fin ( Fig. 7C View Fig ). Posterior to the anal fin, the haemal spines become shorter, more robust, and posteriorly inclined.
MEDIAN FINS. The median fins are not complete. Eight elongate dorsal axonosts are preserved in the holotype, and no dorsal lepidotrichia. The anteriormost axonost is not differentiated relative to more posterior axonosts. Twelve anal axonosts are preserved; again the anteriormost is not differentiated ( Fig. 7B View Fig ). The two forks of the neural and haemal spines insert on either side of an axonost. There is a gap between the axonosts and the lepidotrichia, and a single roundish, extremely weakly ossified baseost is preserved posteriorly. Four basal fulcra are present. The preserved lepidotrichia are unsegmented and are not distally bifurcated. Fringing fulcra are present, with an arrangement similar to that described by Stensiö (1925) for the pelvic fins of S. ornatus , but differing in that the fringing fulcrum and distal lepidotrichium appear to be fused, especially rostrally. When fused, a suture is present between the fulcra and underlying lepidotrichia. The posterior anal fin consists of finer, soft rays. A heavily ossified element associated with more poorly ossified endochondral fragments is also preserved in this region. It is bilaterally symmetrical, forked anteriorly with each of the two rami bifurcating again. The posterior end bears three small projections, with the medial projection being slightly longer than the other two. This element is tentatively interpreted as the axonost plate ( Fig. 7B View Fig ), based on comparisons in shape and position with the same element in Birgeria ( Schwarz 1970) . Caudal radials are preserved, but lepidotrichia are missing.
SQUAMATION. The only scale row present anterior to the median fins is the mid-dorsal scale row. Scales in this row are elongate, needle-like, and unornamented ( Fig. 7C View Fig ). Posterior to the median fins, no scales are present, unlike in S. anningae sp. nov. in which, at minimum, the mid-ventral row is ossified. Immediately anterior to the caudal fin, the caudal peduncle is encased in robust mid-dorsal and midventral scales, which grade into basal fulcra posteriorly ( Fig. 7D View Fig ). The scales in the caudal peduncle are more robust than the abdominal scales, but are also smooth in texture.
Occurrence
Early Jurassic, Toarcian, tenuicostatum Zone , semicelatum Subzone to serpentinum Zone, Baden-Württemberg, Germany; serpentinum Zone, Lower Saxony, Germany; exaratum Subzone , Western Pomerania, Germany; Toarcian, Whitby, UK.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Saurorhynchus hauffi
| Maxwell, Erin E. & Stumpf, Sebastian 2017 |
Belonorhynchus brevirostris
| Woodward A. S. 1895: 18 |
Belonostomus acutus
| Quenstedt 1856: 29 |
Belonorhynchus brevirostris
| Woodward 1899 |
Acidorhynchus brevirostris
| Hauff 1938 |
| Thies (1985 |
Saurorhynchus brevirostris
| Hauff & Hauff 1981 |
| Urlichs et al. (1994) |
| Böttcher (1998) |