
Dysommina orientalis Tighe, Ho & Hatooka, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4454.1.6 |
|
publication LSID |
lsid:zoobank.org:pub:12FBBF69-A222-42AA-A61C-73C0B303B435 |
|
DOI |
https://doi.org/10.5281/zenodo.5976085 |
|
persistent identifier |
https://treatment.plazi.org/id/03B36451-A67D-2346-FF7F-FC19FBE8EA20 |
|
treatment provided by |
Plazi |
|
scientific name |
Dysommina orientalis Tighe, Ho & Hatooka |
| status |
sp. nov. |
Dysommina orientalis Tighe, Ho & Hatooka , sp. nov.
English name: Oriental arrowtooth eel; Japanese name: Suruga-anago
Figs. 1 View FIGURE 1 , 2 View FIGURE 2 , 3A View FIGURE 3 ; Table 1
Dysommina rugosa View in CoL (not of Ginsburg): Hatooka, 1997:8; Chen & Mok, 2001:79; Shao et al., 2008:238; Ho et al., 2015:100.
Holotype: NMMB-P 11131 , 413 mm TL, collected from Dong-gang fishing port, Pingtung, Taiwan, collected by H.- C. Ho, 13 Sep. 2010.
Paratypes (n = 6): USNM 441667 View Materials (1, 316 mm TL; formerly NMMB-P14012 ), Dong-gang fishing port, Pingtung, Taiwan, 20 Oct. 2011 ; NMMB-P3847 (1, 290 mm TL; formerly THUP 4077 View Materials ). Dong-gang fishing port, Pingtung, Taiwan, 21 Mar. 1979 ; NMMB-P8361 (1, 258 mm TL), Dong-gang fishing port, Pingtung, Taiwan, 16 Mar. 2005 ; USNM 441750 View Materials (1, 325 mm TL; formerly NSYSU 3028 ), Dong-gang fishing port, Pingtung, Taiwan, Jan. 1996 ; ASIZP 57954 View Materials (1, 238 mm TL), Dong-sha Island, South China Sea, 17 Aug. 1991 ; OMNH-P10000 (1, 625 mm TL), Suruga Bay , Shizuoka Prefecture, Japan, 34°54’N, 138°30.5’E, 300–400 m depth, 17 Dec. 1996. GoogleMaps
Diagnosis. A species of the genus Dysommina with the following combination of characters: predorsal vertebrae 11–12, total vertebrae 137–141, eye diameter less than 10% head length, vomerine dentition reduced to 3 large compound teeth set in papillose pads with a fourth small tooth more posteriorly, maxillary and mandibular teeth numerous but small.
Description. Body moderately elongate, slightly compressed in head and trunk. Pectoral fin present; dorsal-fin origin over tip of pectoral fin; anal fin origin more than 1 head length behind tip of pectoral fin. Snout projects slightly beyond tip of lower jaw. Body light greyish-brown dorsally, paler ventrally; dorsal and anal fins with light margins; posterior one third of anal-fin base and posterior one-seventh of anal fin dark.
Morphometric data of the holotype (in mm): total length 413; predorsal length 65.4; preanal length 119; tail length 294; head length 51.2; body depth at gill opening 19.6; body depth at anus 24.6; eye diameter 4.1; interorbital width 8.0; snout length 13.3; upper jaw length 24.4; lower jaw length 23.5; pectoral fin length 10.5.
Proportions as percentage of total length: predorsal length 15.8 (14.0– 16.2 in paratypes); preanal length 30.0 (27.0–29.8); tail length 70.0 (70.2–73.0); head length 12.8 (12.6–15.0); body depth at gill opening 5.6 (4.9–5.3); body depth at anus 5.8 (3.7–5.9). Proportions as percentage of head length: eye diameter 8.5 (7.6–9.8); interorbital width 16.0 (17.1–19.5); snout length 25.1 (23.2–27.8); upper jaw length 48.1 (43.9–48.0); lower jaw length 44.3 (39.0–43.4). Meristic data for the holotype (paratypes in parentheses): total vertebrae 139 (137–141); predorsal vertebrae 11 (11–12); preanal vertebrae 30 (26–32); dorsal rays 310 (304–321); anal rays 295 (260–289); anal origin at dorsal ray 50 (51–60).
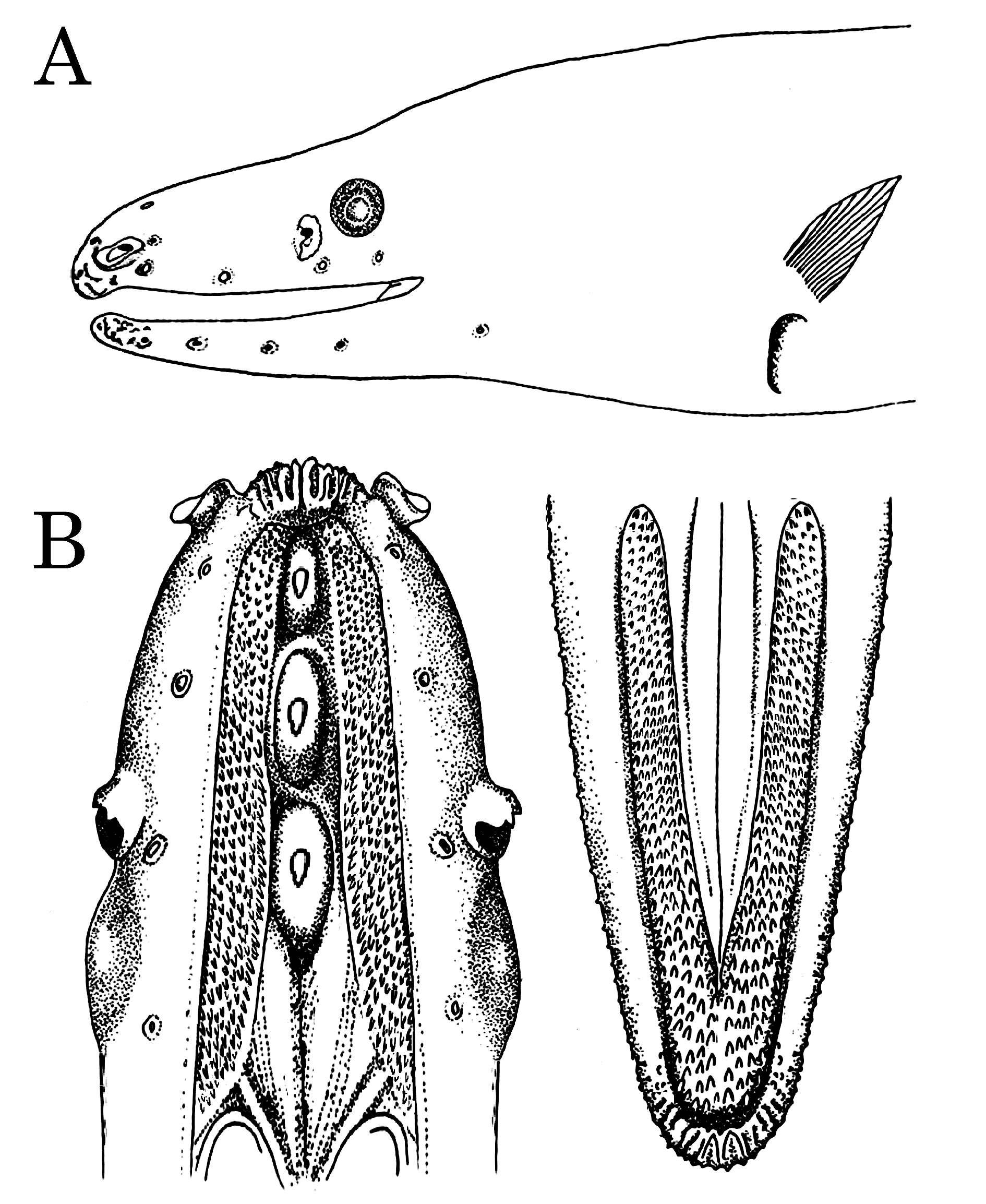
Cephalic lateralis pores ( Fig. 2A View FIGURE 2 ): supraorbital 3 (3); adnasal 1 (1); infraorbital 4 (4); preoperculomandibular 6 (6). Lateral line pores absent.
Dentition ( Fig. 2B View FIGURE 2 ): Intermaxillary teeth absent. Three relatively large, compound vomerine teeth set in papillose pads; usually a much smaller compound tooth posteriorly on the palate. Mandibular teeth set in a band composed of 5–7 irregular rows increasing in size gradually from outer to inner with approximately 44–50 teeth in the inner row; maxillary teeth similar, but set in a band composed of 6–8 irregular rows increasing in size gradually from outer to inner with approximately 50–60 teeth in the inner row. Bands of both mandibular and maxillary teeth extend well back along the jaws.
Distribution. Known from the type specimens collected from the South China Sea off Taiwan and Dongsha Islands, and from Suruga Bay, Japan. This species is probably more widespread in the western Pacific. The bathymetric range is estimated to be 200–400 m based on the collection data of one paratype and other fishes collected together.
Etymology. The name is derived from the Latin orientalis , “of the east” and refers to the type locality in the Far East off Taiwan and Japan.
Comparison. Morphometric and meristic data for the holotype and study material of Dysommina rugosa and the type series of Dysommina orientalis are given in Table 1. One paratype, ASIZP 57954, is excluded from this comparison because it has a regenerated tail which biases all of the morphometric and meristic data.
Dysomma orientalis is very similar in overall appearance to D. rugosa , but differs in several key characters. The first character is the shorter predorsal length in D. orientalis (14.0–16.2 % TL versus 16.1–21.1 in D. rugosa ). Related to the shorter predorsal length is a lower number of predorsal vertebrae (11–12 versus 14–15) and a higher number of dorsal rays anterior to the anal origin (50–60 versus 38–48).
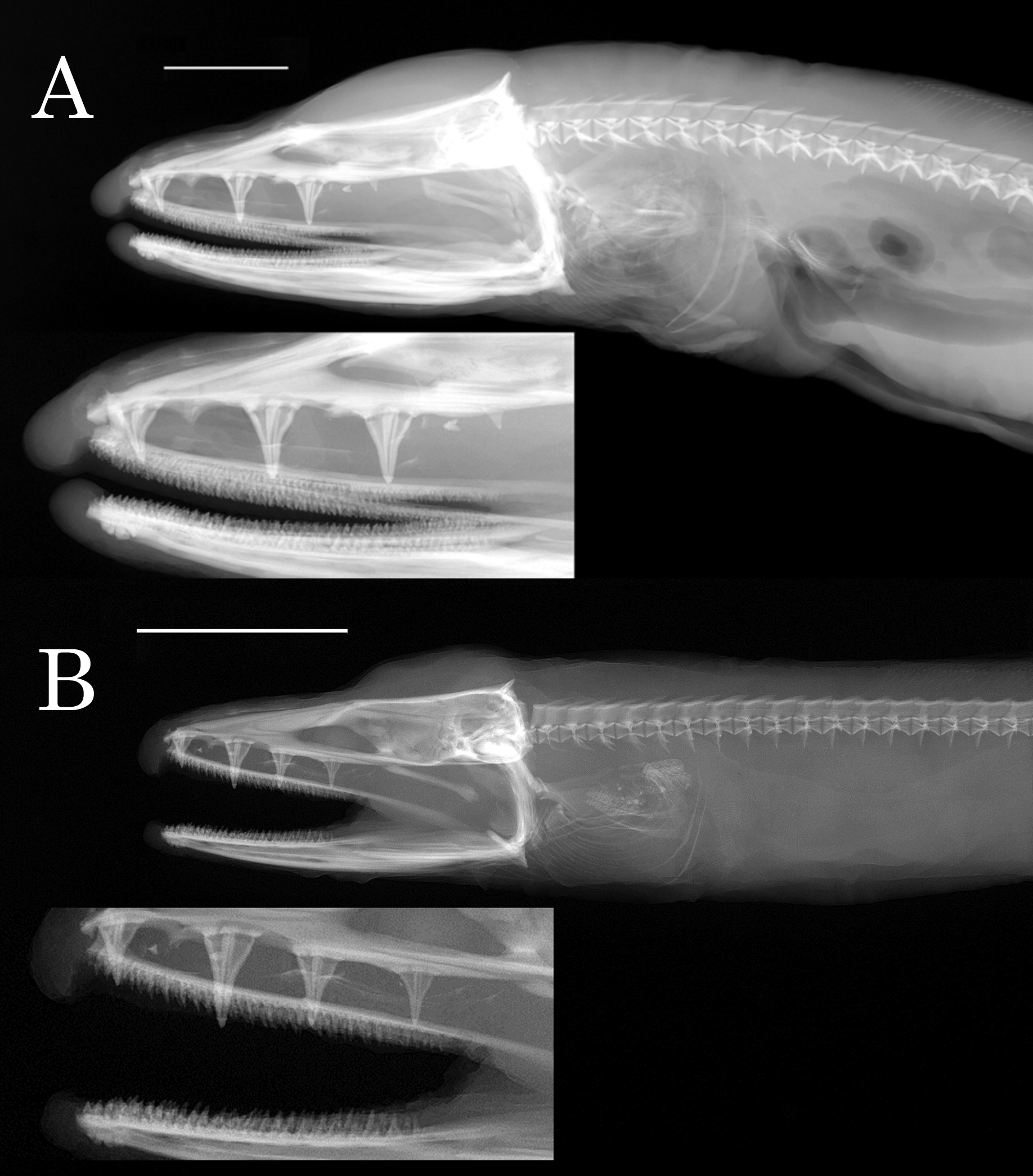
The second character distinguishing D. orientalis from D. rugosa is the dentition. The number of large, compound teeth set in pads of papillose tissue is less in D. orientalis (3 versus 4). The position of the teeth along the vomer is also different. Figure 3 View FIGURE 3 shows x-rays of the skulls of the holotype of D. orientalis and one of the comparative specimens of D. rugosa . In D. orientalis , the first tooth is set slightly back from the tip of the ethmovomer, the second is approximately halfway between the tip of the ethmo-vomer and the posterior margin of the orbit, and the third is set near the posterior third of the orbit. In D. rugosa , the first tooth is set near the tip of the ethmo-vomer, the second is approximately halfway between the tip of the ethmo-vomer and the anterior margin of the orbit, the third is set near the anterior margin of the orbit, and the fourth is set under the center of the orbit. In addition to the differences in the large vomerine teeth, most of our D. orientalis has a fourth small vomerine tooth set well back on the palate behind the orbit. D. rugosa does not have a corresponding tooth. There are 2 specimens, one of each species, that have an extra vomerine tooth. However, in both of these specimens, there are 2 close-set teeth that are in the same pad of papillose tissue. These pairs of teeth probably represent an original tooth that was injured and a subsequent replacement tooth. The original tooth was not injured enough to break off, resulting in both the original tooth and the replacement tooth together in a single pad.
In addition to the differences in the vomerine dentition, there are significant differences in the mandibular and maxillary dentition. The number of tooth rows in the bands of teeth on both the mandible and the maxilla are more abundant in D. orientalis than in D. rugosa . There are 5–7 tooth rows across the mandibular tooth band in D. orientalis and 3–5 in D. rugosa . Moreover, there are 44–50 teeth in the inner mandibular row in D. orientalis and only 20–30 in D. rugosa . In the maxillary tooth bands, there are 6–8 tooth rows in D. orientalis and 4–5 in D. rugosa , and there are 50–60 teeth in the inner maxillary row versus 24–42 in in D. rugosa . In D. orientalis , both the mandibular and maxillary tooth bands extend further posteriorly than in D. rugosa .
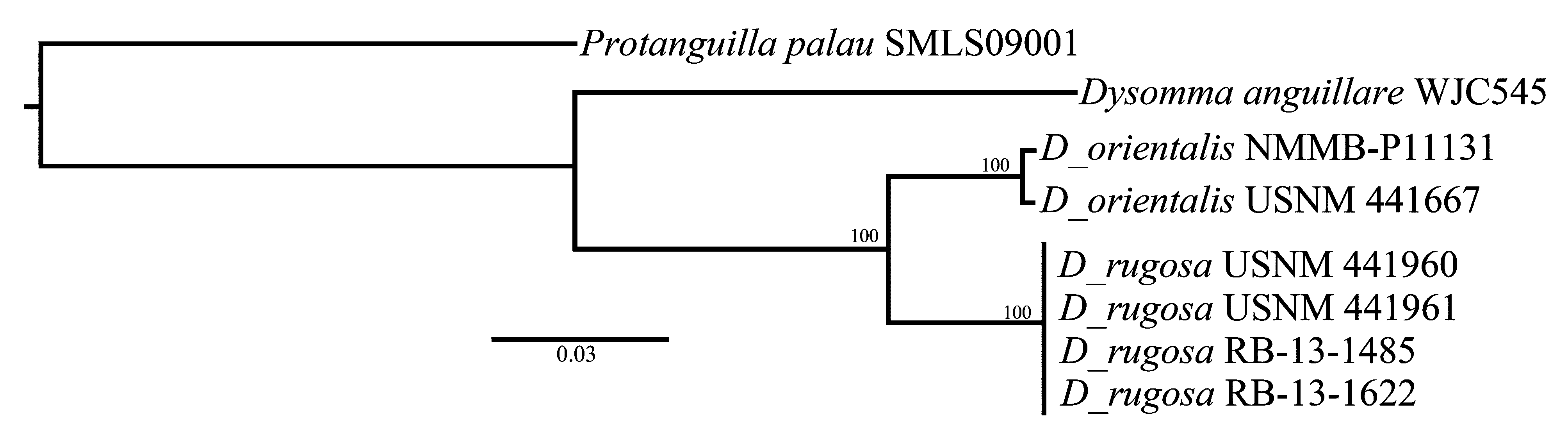
The final and conclusive character that distinguishes D. orientalis and D. rugosa is the DNA sequences. Table 2 shows the distance matrices for COI, 16S and Combined. As can be seen in the table, the COI sequences for the four samples of D. rugosa were identical, and 7.2–7.3 % different from the two samples of D. orientalis . The 16S sequences for the four samples of D. rugosa were also identical and differed 0.8–1.0 % from the two samples of D. orientalis . The differences between the two species for the combined COI and 16S sequences ranged from 4.1 to 4.5 %, clearly indicating that they are two distinct species. Figure 4 View FIGURE 4 is a maximum likelihood tree of combined COI and 16S sequences showing the difference between the 2 species compared to the two outgroups, Dysomma anguillare and Protanguilla palau . The high bootstrap value for the node linking D. orientalis and D. rugosa indicates the close relationship between the two species showing that they belong together in the genus Dysommina .
Remarks. The present species has long been recognized as Dysommina rugosa , a species originally described from the western Atlantic Ocean. Specimens collected from Taiwan and Japan are now described as D. orientalis . The records from Hawaii ( Robins & Robins 1976) and specimens collected from the Solomon Islands, Philippines, Madagascar and New Caledonia (listed in Ho et al. 2015) as well as the specimens reported from Vailulu'u Seamount ( Staudigel et al. 2006) are presently under study by the senior author and apparently represent several cryptic species in the genus Dysommina . A redescription of D. rugosa sensu stricto is being prepared by the senior author and will discuss the variation of this species as well as its geographic and bathymetric distribution.
Ho et al. (2015) also pointed out that some members of the genus Dysomma have multiple rows of teeth on both jaws and lacking intermaxillary teeth which may suggest they are congeners of Dysommina rather than Dysomma . The osteology and relationships of members of the subfamily Ilyophinae are under study by the senior author and will probably result in several changes in the generic taxonomy of these eels.
Comparative materials. Dysommina rugosa : USNM 131594 (holotype, 196 mm TL), off Cumberland Island, Georgia, United States, Atlantic Ocean, 30°53’00”N, 79°42’30”W, 499 m, 5 May 1886. USNM 44324 (1, 283 mm TL), off Savannah, Georgia, United States, Atlantic Ocean, 31°09’00”N, 79°33’30”W, 644 m, 5 May 1886. USNM 179213 (1, 194 mm TL), off Florida, Gulf of Mexico, 28°16’N, 85°50’W, 439 m, 3 Dec 1962. USNM 190541 (1, 265 mm TL), Southwest of Dry Tortugas, Florida Keys, Gulf of Mexico, 24°28’N, 83°24’W, 329 m, 7 Jun 1959. USNM 200776 (1, 248 mm TL), East coast of Florida, 29°59’N, 80°09’W, 347 m, 21 Nov. 1965. Atlantic Ocean, 404.2 m, 6 Aug. 2009. USNM 441960 (1, 226 mm TL), USNM 441961 (1, 224 mm TL), off Maryland, Baltimore Canyon seep, Atlantic Ocean, 38°02’53”N, 73°49’19”W, 398 m, 16 May 2013. USNM 441962 (1), off Maryland, Baltimore Canyon seep, 414 m, 7 Sep. 2012. To be deposited at North Carolina Museum of Natural Sciences: CH- 06-018 (1, 174 mm TL); JSL-05-4894 (1, 128 mm TL); JSL-4362 (1, 184 mm TL); JSL-4364 (1, 176 mm TL); JSL-4366 (3, 145- 217 mm TL); JSL-4894 (2, 128- 157 mm TL); ROV-2012-NF-14 (1, 193 mm TL); ROV-2013- RB-689 (2, 220- 225 mm TL); SJ-02-036 (1, 210 mm TL); SJ-2004-025 (1, 96 mm TL).

| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Dysommina orientalis Tighe, Ho & Hatooka
| Tighe, Kenneth A., Ho, Hsuan-Ching & Hatooka, Kiyotaka 2018 |
Dysommina rugosa
| Hatooka, 1997 :8 |
| Chen & Mok, 2001 :79 |
| Shao et al., 2008 :238 |
| Ho et al., 2015 :100 |