
Monoctenus obscuratus hokkaidonis Hara, 2023
|
publication ID |
https://doi.org/10.11646/zootaxa.5380.6.3 |
|
publication LSID |
lsid:zoobank.org:pub:AA915DFA-58B9-428D-AF5D-193E3295AB04 |
|
DOI |
https://doi.org/10.5281/zenodo.10371571 |
|
persistent identifier |
https://treatment.plazi.org/id/03B38786-FF84-CD4C-FF21-FF54FB24574E |
|
treatment provided by |
Plazi |
|
scientific name |
Monoctenus obscuratus hokkaidonis Hara |
| status |
subsp. nov. |
Monoctenus obscuratus hokkaidonis Hara , subsp. nov.
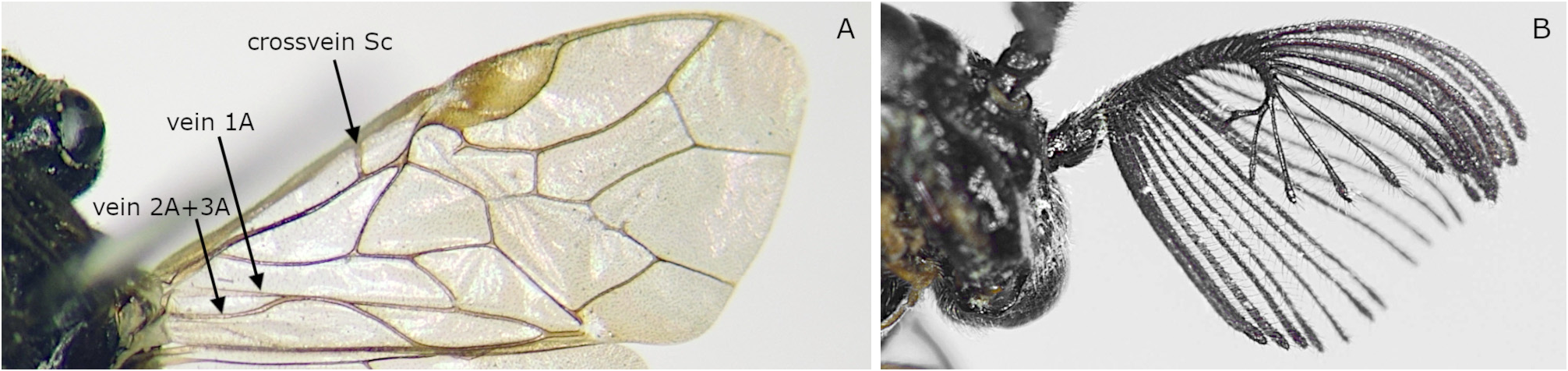
( Figs 3A View FIGURE 3 2, B2b View FIGURE 2 , 4A 2 View FIGURE 2 , 9A 2 View FIGURE 2 , 20, 21)
Description. Female. Length 4.5–6.0 mm (6.0 mm in holotype). Black ( Fig. 20A, B View FIGURE 20 ). Mandible red brown on apical half. Palpi yellow, basally darkened. Pronotum black ( Fig. 3B View FIGURE 3 2b View FIGURE 2 ), sometimes narrowly dark brown on posterodorsal corner. Tegula sometimes laterally dark brown. Legs yellow on narrow apices of coxae, apices of femora, and tibiae and tarsi ( Fig. 20B View FIGURE 20 ); tarsi apically brown or dark brown; hind tibia often narrowly yellow brown apically. Wings clear transparent, very slightly brownish ( Fig. 20A View FIGURE 20 ); veins dark brown, partly pale brown; stigma brown, apically widely pale brown.
Body surface mostly shiny and smooth; punctures indistinct or minute.
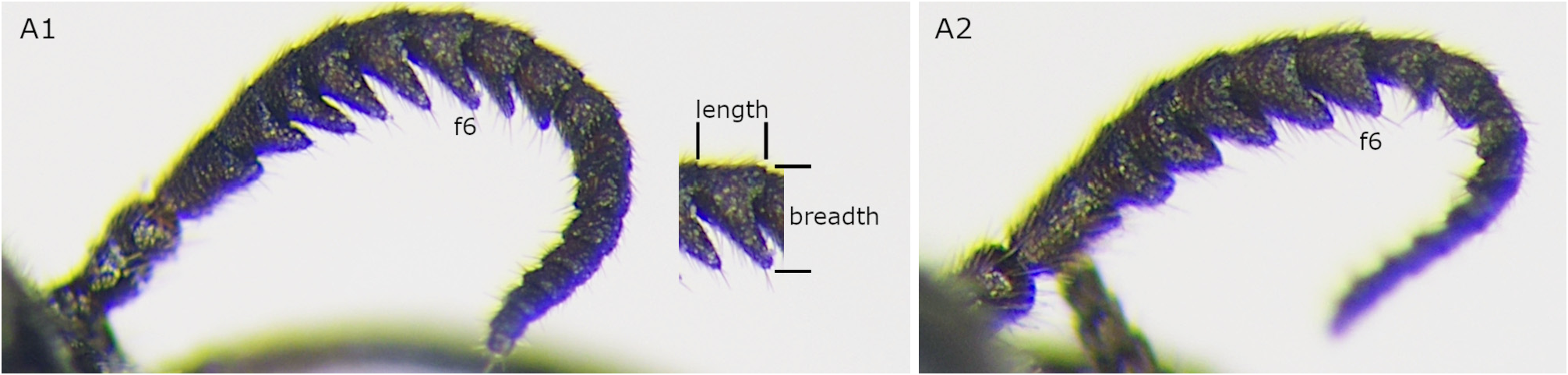
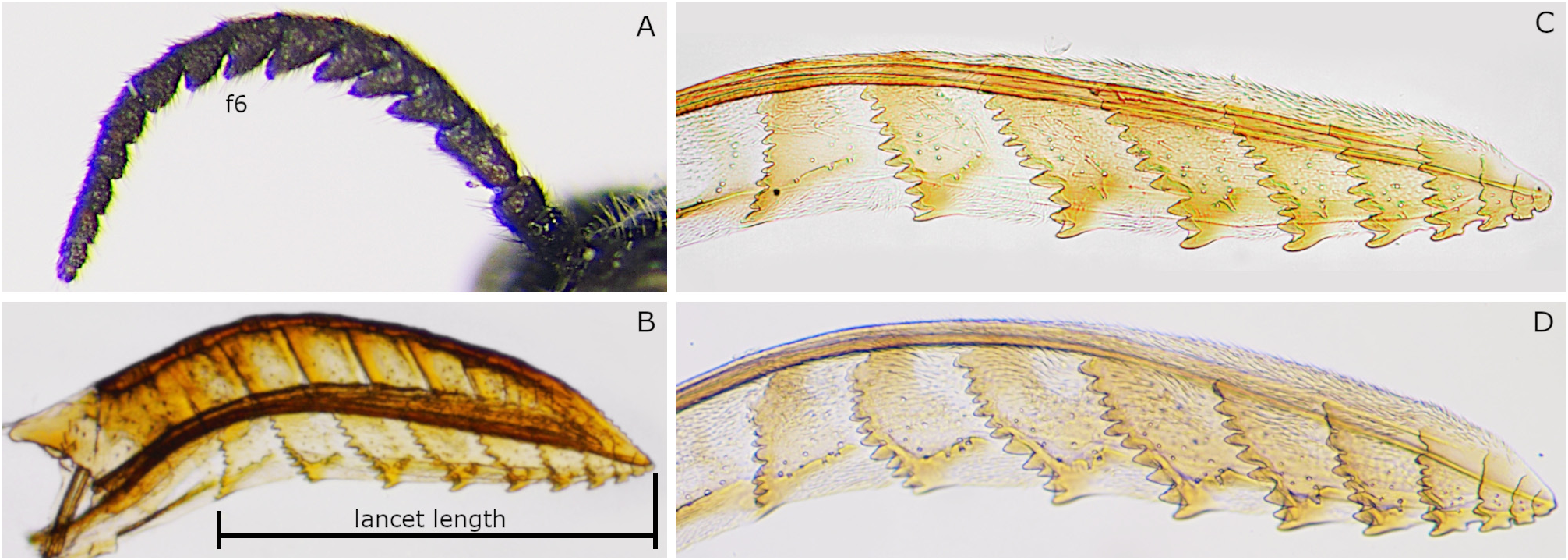
Clypeus with median furrow dorsally; ventral edge concave medially. Antenna with 18 antennomeres ( Figs 4A View FIGURE 4 2 View FIGURE 2 , 20E); flagellum serrations short; flagellomere 6 in lateral view with breadth including serration 1.1–1.4 × dorsal length ( Figs 4A View FIGURE 4 2 View FIGURE 2 , 20E). Valvula 3 apically slightly concave in dorsal view ( Fig. 20F View FIGURE 20 ), apically truncate in lateral view ( Fig. 20G View FIGURE 20 ); apical groove rather wide ( Fig. 20H View FIGURE 20 ). Lance in lateral view with dorsal edge not or slightly concave basally ( Fig. 20I, J View FIGURE 20 ). Lancet with 10 annuli and apex acute ( Fig. 6K–M View FIGURE 6 ); length from ventral end of ctenidium 1 to apex 3.7–4.1 × breadth; ctenidia 1 and 2 converging dorsally; ctenidia 2–6 inclined anteriorly.
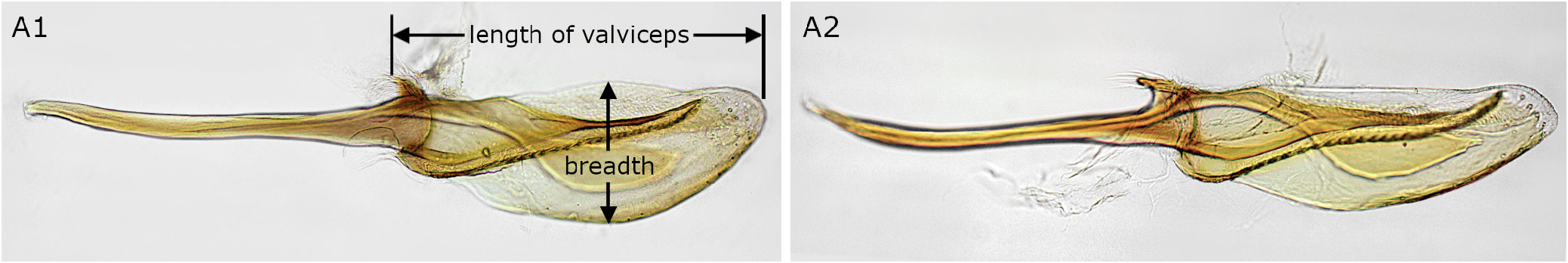
Male. Length 4.5–6.0 mm. As in female except for usual sexual differences ( Fig. 20C, D View FIGURE 20 ). Antenna with 23–24 rami (26–27 antennomeres). Subgenital plate in ventral view truncate apically. Genitalia Fig. 20N View FIGURE 20 ; in penis valve, paravalva with ventral edge nearly straight and inner sclerite widest near middle ( Figs 9A View FIGURE 9 2 View FIGURE 2 , 20O); valviceps 3.2–3.5 × as long as broad ( Figs 9A View FIGURE 9 2 View FIGURE 2 , 20O).
Immature stages. Middle instar larva ( Fig. 21B View FIGURE 21 ): head brown; trunk yellow; thoracic legs gray. Late instar larva ( Fig. 21C View FIGURE 21 ): 13 mm long; head brown, dorsolaterally dark brown, anteriorly yellow brown; trunk pale yellow green, with narrow gray middorsal and supraspiracular longitudinal stripes; thoracic legs gray. Cocoon: 8 mm long in female, 6.5 mm long in male; dark brown; double-walled, with outer wall loosely spun and inner wall tightly spun.
Material examined. Holotype ( Figs 3A View FIGURE 3 2, B2b View FIGURE 2 , 20A, B, E–I, K, 21C): ♀, labelled “ JAPAN, Hokkaido, Fu- kagawa, Takadomari , em. 11. V. 2010, H. HARA ”, “ HH090705 G, Larva coll. 5. VII. 2009, coc. 13. VIII., Host: Juniperus communis var. montana ” . Paratypes: 1♂, same locality and collector as holotype except 26. V. 2007 ( Fig. 9A View FIGURE 9 2 View FIGURE 2 ) ; 2♀, ditto except 26. V. – 6. VI. 2007, by Malaise trap ( Figs 4A View FIGURE 4 2 View FIGURE 2 , 20J, L, M) ; 1♂, same locality and collector as holotype except coll. solitary larva on Juniperus communis var. montana 3. VII. 2010, coc. late VII – late VIII., em. 12 IV. 2011 ( Fig. 20C, D, N, O View FIGURE 20 ) .
Etymology. The name hokkaidonis is derived from the type locality and is to be treated as an adjective.
Distribution. Japan ( Hokkaido).
The nominotypical subspecies is known from Europe (Hartig 1937, Taeger et al. 2018) and Siberia ( Sundukov 2017b).
Host plant. Cupressaceae : Juniperus communis L. var. montana Aiton.
The hosts of the nominotypical subspecies are J. communis L. subsp. communis , J. communis L. subsp. alpina (Suter) Čelak ( Viitasaari & Varama 1987) .
Life history. The adults collected from late May to early June. The eggs ( Fig. 21A View FIGURE 21 ) were observed in middle June. They were singly inserted in current-year needles, but about 20 eggs were observed on adjacent new shoots. The solitary middle-instar larvae (about 9 mm long) were collected in early July ( Fig. 21B View FIGURE 21 ) and they became cocoons in early to late August. The adults emerged in the following spring. This sawfly has one generation per year.
Remarks. Monoctenus obscuratus (Hartig, 1837) has been characterized in having an almost entirely black body, mostly blackish brown tarsi and hind tibia and distinctly infuscated wings (see Hartig 1837, Enslin 2017, Gussakovskij 1947, Viitasaari & Varama 1987, Zhelokhovtsev & Zinovjev 1988, Mol & Aarsten 1994, Lacourt 2020; images of the female and male syntypes of Lophyrus obscuratus in Taeger et al. 2018), although Viitasaari & Varama (1987) stated that the wings of M. obscuratus were usually darker, but the darkness was unreliable as a character. However, Zombori (1975) reported the specimens with an entirely yellow hind tibia under the name of M. obscuratus intermedius , and Borowski & Stawski (2021) did not mention dark legs and dark wings as characters of M. obscuratus , and their photos of a female and male of M. obscuratus have an entirely yellow hind tibia and slightly infuscated wings (fig. 226 in Borowski & Stawski 2021). The Hokkaido specimens ( Fig. 20A–D View FIGURE 20 ) are very similar to fig. 226 in Borowski & Stawski (2021), and the valviceps of the Hokkaido males ( Figs 9A View FIGURE 9 2 View FIGURE 2 , 20O) are elongated with the paravalva very thin and pointed, just like that of M. obscuratus (fig. 12b in Viitasaari & Varama 1987, fig. 4 in Mol & Aarsten 1994). Therefore, the Hokkaido specimens are considered to belong to M. obscuratus .
The Hokkaido specimens slightly differs from the European specimens. The Hokkaido males have an antenna with 23–24 rami (26–27 antennomeres) and a valviceps 3.2–3.5 × as long as broad ( Figs 9A View FIGURE 9 2 View FIGURE 2 , 20O), but the European males have an antenna with 22 rami and 25 antennomeres ( Enslin 1917, Gussakovskij 1947, Viitasaari & Varama 1987, Zhelokhovtsev & Zinovjev 1988, Mol & Aarsten 1994) and a valviceps 2.9–3.1 × as long as broad (fig. 12b in Viitasaari & Varama 1987, fig. 4 in Mol & Aarsten 1994). The Hokkaido larva has a pale yellow green trunk with narrow black longitudinal stripes ( Fig. 21C View FIGURE 21 ), but the European larva has yellow-gray or greenish with broad black longitudinal stripes according to Viitasaari & Varama (1987, photo in p. 37) or a pink-purple trunk without clearly marked longitudinal stripes according to Borowski & Stawski (2021). Therefore, we believe that the Hokkaido specimens represent a separate subspecies.
Monoctenus obscuratus hokkaidonis is very similar to M. nipponicus but distinguished from it by the characters mentioned in the key and the remarks of M. nipponicu s. These two taxa also have slightly different lancets. The lancet of the former ( Fig. 20K–M View FIGURE 20 ) is slightly narrower than that of the latter ( Fig. 19K, L View FIGURE 19 ); the lancet length from the ventral end of ctenidium 1 to the apex is 3.7–4.1 × the lancet breadth in M. obscuratus hokkaidonis but 3.4–3.6 × in M. nipponicus . This subspecies is also similar to M. juniperi and M. kondoi . Their differences are as follows. The abdomen of the former female is almost completely black ( Figs 3A View FIGURE 3 2 View FIGURE 2 ), but those of the latter two females have yellow laterotergites ( Figs 3A View FIGURE 3 1 View FIGURE 1 ). The lancet of M. obscuratus hokkaidonis ( Fig. 20K–M View FIGURE 20 ) is slightly broader than that of M. juniperi ( Fig. 18C, D View FIGURE 18 ) and slightly narrower than that of M. kondoi ( Fig. 17G View FIGURE 17 ); the lancet length from the ventral end of ctenidium 1 to the apex is 3.7–4.1 × the lancet breadth in M. obscuratus hokkaidonis , 4.4–4.6 × in M. juniperi and 3.2 × in M. kondoi . The number of male antennal rami is 23–24 in M. obscuratus hokkaidonis but 17–21 in M. juniperi ( Enslin 1917, Gussakovskij 1947, Viitasaari & Varama 1987, Zhelokhovtsev & Zinovjev 1988, Mol & Aarsten 1994) and 26 in M. kondoi . The penis valve of M. obscuratus hokkaidonis also differs from those of M. juniperi and M. kondoi ; the valviceps has the length 3.2–3.5 × the breadth in M. obscuratus hokkaidonis ( Figs 9A View FIGURE 9 2 View FIGURE 2 , 20O), 2.3–2.7 × in M. juniperi (fig. 12a in Viitasaari & Varama 1987, fig. 6 in Mol & Aarsten 1994) and 2.5 × in M. kondoi ( Fig. 17H, I View FIGURE 17 ). For more differences between M. obscuratus hokkaidonis and M. kondoi , see the key.
Monoctenus obscuratus hokkaidonis is separated from other species of Monoctenus by the coloration (compare with the descriptions of other species of the genus by Norton 1872, Cresson 1880, Marlatt 1888, Gussakovskij 1947, Smith 1975, Smith et al. 2010, De Lira-Ramos et al. 2022, Japoshvili & Haris 2022, this study).







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |