
Williamsrhizoecus udzungwensis Schneider & LaPolla, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4853.2.9 |
|
publication LSID |
lsid:zoobank.org:pub:DEAF3CA3-6B57-4695-9D97-6838C2103810 |
|
DOI |
https://doi.org/10.5281/zenodo.4410839 |
|
persistent identifier |
https://treatment.plazi.org/id/03B387A5-B91B-FFFF-FF43-F88D3DEBFDA7 |
|
treatment provided by |
Plazi |
|
scientific name |
Williamsrhizoecus udzungwensis Schneider & LaPolla |
| status |
sp. nov. |
Williamsrhizoecus udzungwensis Schneider & LaPolla sp. n.
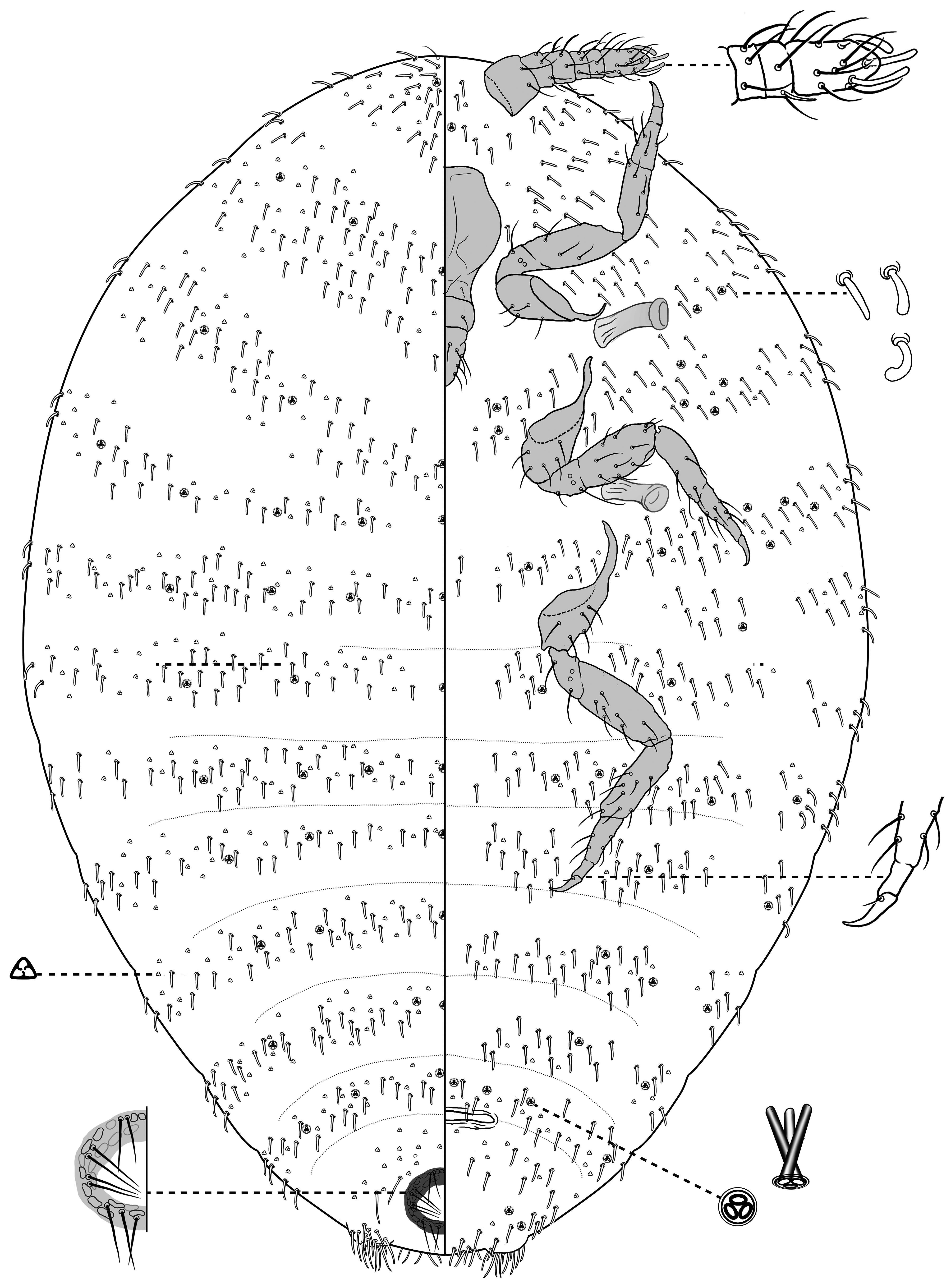
Fig. 1 View FIGURE 1
Material examined. Holotype adult female: TANZANIA: Udzungwa Mountains , attended by / colony of Acropyga silvestrii within nest / chambers, under rocks alongside a stream, / -7.8449, 36.8835, 350m; 27 March 2011; / J.S. LaPolla coll. (JSL110327-04A) ( USNM). GoogleMaps
Paratypes: TANZANIA: 1 adult female, same data as holotype (JSL110327-02A) ( USNM) GoogleMaps ; 1 adult female, same data as holotype (JSL110327-02B) ( USNM) GoogleMaps ; 1 adult female, same data as holotype (JSL110327-03A) ( USNM) GoogleMaps ; 1 adult female, same data as holotype (JSL110327-03B) ( USNM) (voucher ID: S0430A) GoogleMaps ; 2 immature instars together on 1 slide, same data as holotype (JSL110327-03C) ( USNM) GoogleMaps ; and 1 adult female, same data as holotype (JSL110327-04B) ( USNM) GoogleMaps .
Description of adult female (N = 6). Mounted on microscope slide, body approximately oval and membranous throughout, 0.88–0.95 mm long, 0.64–0.73 mm wide, widest near abdominal segment II. Abdomen gently tapering toward posterior end; abdominal segment VIII approximately 250 μm wide at base. Anal lobes poorly developed, indicated by slight protrusions of the body margin, each lobe with a cluster of several dorsal clavate setae, each seta 10–18 μm long. Anal ring dorsal, located slightly anterior of body apex, with two concentric rows of ovoid cells, some lateral cells each bearing a spicule. Anal ring setae arranged in 9 pairs, numbering 18 in total, each seta approximately 19–24 μm long. Antennae 6-segmented, situated close to each other on ventral submargin of head. Stout flagellate setae present on each antennal segment, 12–21 μm long; with 1 falcate sensory seta present on segment V and 3 falcate sensory setae present on segment VI. One sensory pore and two seta-like sensillae present on antennal segment II. Average antennal segment lengths in μm: I – 32, II – 19, III – 16, IV – 12, V – 13, VI – 38; overall length 130 μm. Eyes absent. Dorsal ostioles absent.
Labium 3-segmented, 76 μm long and 48 μm wide, with few short flagellate setae present on each segment. Cephalic plate absent. Legs well developed; hind leg average length measurements in μm: trochanter + femur 108; tibia + tarsus 129. Ratio of lengths of metatrochanter + femur to tibia + tarsus, 0.84; ratio of lengths of metatibia to tarsus, 1.39. Hind tarsus widest at base and tapering to claw, about 24 μm long. Claws with simple digitules, shorter than the claw, and without denticles. Several flagellate setae present on each leg segment, about 15 setae present on each hind tibia. Spiracles normal for the family, each approximately 23 μm in diameter at widest point. Circulus absent.
All body setae on dorsum and venter clavate to falcate, 8.5–10 μm long, either linear or curved, widest at tip or widest just proximal to tip and tapering distally. Setae arranged in irregular rows separated by bald intersegmental regions, otherwise fairly evenly distributed throughout. Trilocular pores numerous across dorsum and venter, interspersed with setae. Tritubular pores also numerous on both dorsum and venter, though less common than trilocular pores; arranged in singular transverse rows toward the midline or posterior edge of each segment, those on dorsum of abdomen mostly located medially to submarginally, those on venter of abdomen mostly located marginally to submarginally, except for segment VII, where they extend from midline to margin. Multilocular disc pores absent.
Comments. Williamsrhizoecus udzungwensis sp. n. is similar to W. coffeae in that they both possess 6 antennal segments, lack ventral abdominal circuli, and have clavate body setae. However, in W. udzungwensis sp. n. ( Fig. 1 View FIGURE 1 ) the body setae are short and thickened throughout the length of the seta, sometimes curved and appearing falcate (sickle-shaped), whereas in W. coffeae the body setae have a longer flagellate stem that terminates in a dilated tip. The same trait distinguishes W. udzungwensis sp. n. from W. baskyi , which is also from Tanzania. Additionally, whereas W. udzungwensis sp. n. lacks circuli and has six antennal segments, W. baskyi has two circuli and only five antennal segments.
There are similarities between W. udzungwensis sp. n. and the only other rhizoecid confirmed to associate with Acropyga in the Old World ( Tanaka 2016), Ishigakicoccus shimadai Tanaka. Besides the obvious ecological connection, both species lack dorsal ostioles and both have body setae that could be described as clavate. The body setae of I. shimadai are mostly flagellate and hooked at the apex, but rarely they are knobbed at the apex ( Tanaka 2016), suggesting a potential affinity to Williamsrhizoecus . Tanaka considered Ishigakicoccus as being similar to Capitisetella and Pseudorhizoecus because they all lack dorsal ostioles, but did not consider Williamsrhizoecus in his discussion, which differs from the other genera by having tritubular pores. Tanaka used the presence of two types of wax pores in Ishigakicoccus , small pores with 6 loculi surrounding a central chamber and large 3–5 locular pores without a central chamber, as justification for establishing a new genus ( Tanaka 2016). These structures are distinct from the multilocular disc pores found in Williamsrhizoecus , which are present only in W. coffeae .
Acropyga workers were observed actively carrying individuals of W. udzungwensis sp. n. around within the nest-box and gathering them together into small chambers that the workers had excavated from loose soil. The behavior of arranging root mealybugs into protected clusters is a critical observation of direct association. No mating swarms of A. silvestrii were observed, so trophophoresy of W. udzungwensis sp. n. (transportation of gravid females by A. silvestrii queens) cannot be confirmed at present.
Prior to this study, the only known root mealybug association with an African Acropyga species was between A. arnoldi Santschi and the xenococcid, Eumyrmococcus scorpioides (De Lotto) ( Prins 1982; LaPolla & Spearman 2007). Eumyrmococcus williamsi Kozár & Konczné Benedicty also occurs in Tanzania, in the Uluguru Mountains, but the Acropyga species associated with this xenococcid was not recorded at the time of its collection ( Kozár & Konczné Benedicty 2007). Eumyrmococcus williamsi was later found to associate with a new species of Acropyga (JSL manuscript in preparation). Interestingly, the neighboring Udzungwa and Uluguru Mountain ranges each harbor a distinct Acropyga species partnered with a root mealybug.
Ecological associates. Acropyga silvestrii Emery ( Hymenoptera : Formicidae ); W. udzungwensis sp. n. was feeding on roots, host plant not recorded.
Etymology. The specific epithet is an adjective formed from Udzungwa, the mountain range where it was discovered, together with the Latin suffix - ensis, meaning of or from a place.
| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |