
Caliroa oishii ( Takeuchi, 1933 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4768.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:C8036F69-F881-4727-96E7-C78AA6C7F920 |
|
DOI |
https://doi.org/10.5281/zenodo.3794899 |
|
persistent identifier |
https://treatment.plazi.org/id/03B387A9-FFDF-FF89-1DC6-2450314DF8C9 |
|
treatment provided by |
Plazi |
|
scientific name |
Caliroa oishii ( Takeuchi, 1933 ) |
| status |
|
Caliroa oishii ( Takeuchi, 1933)
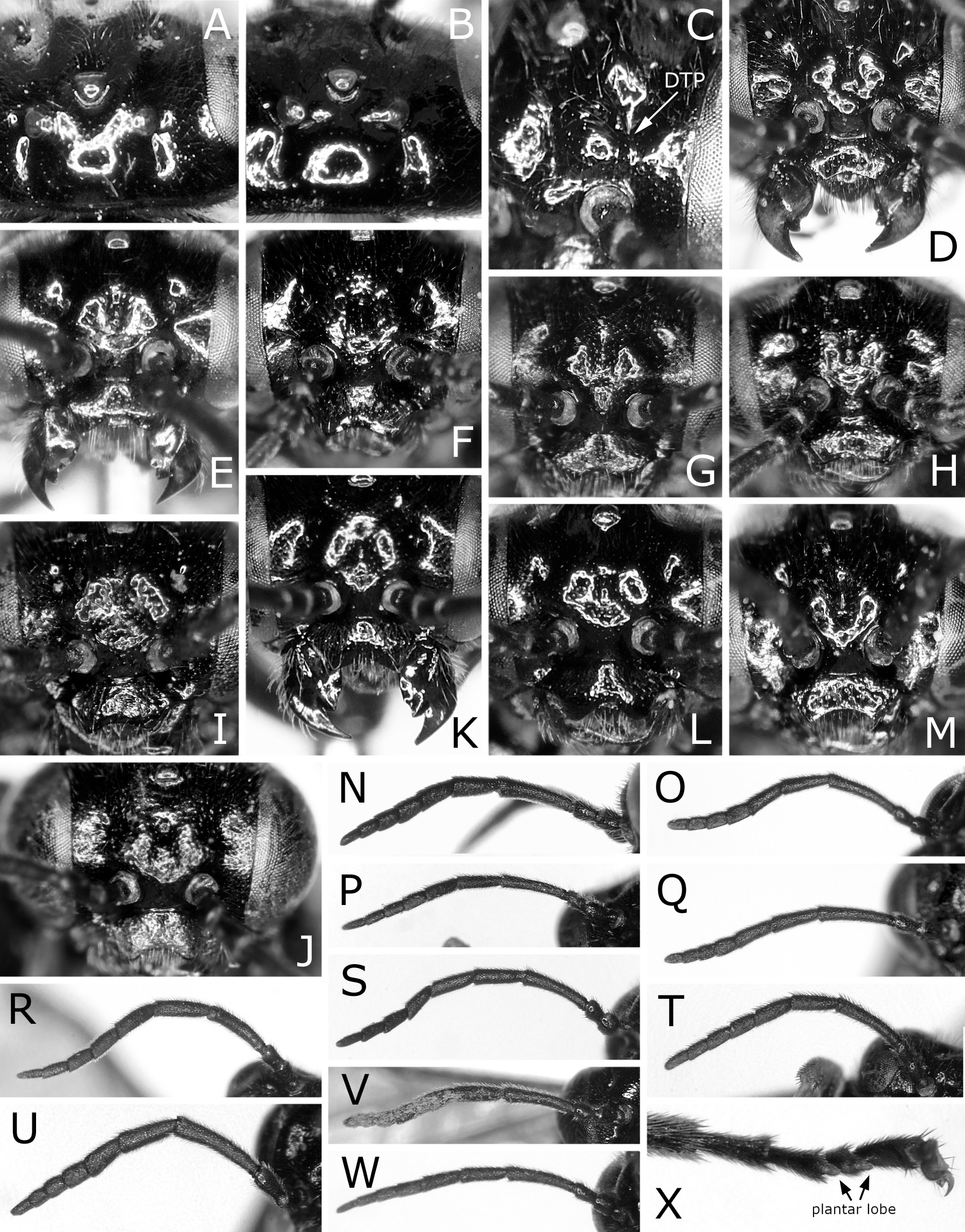
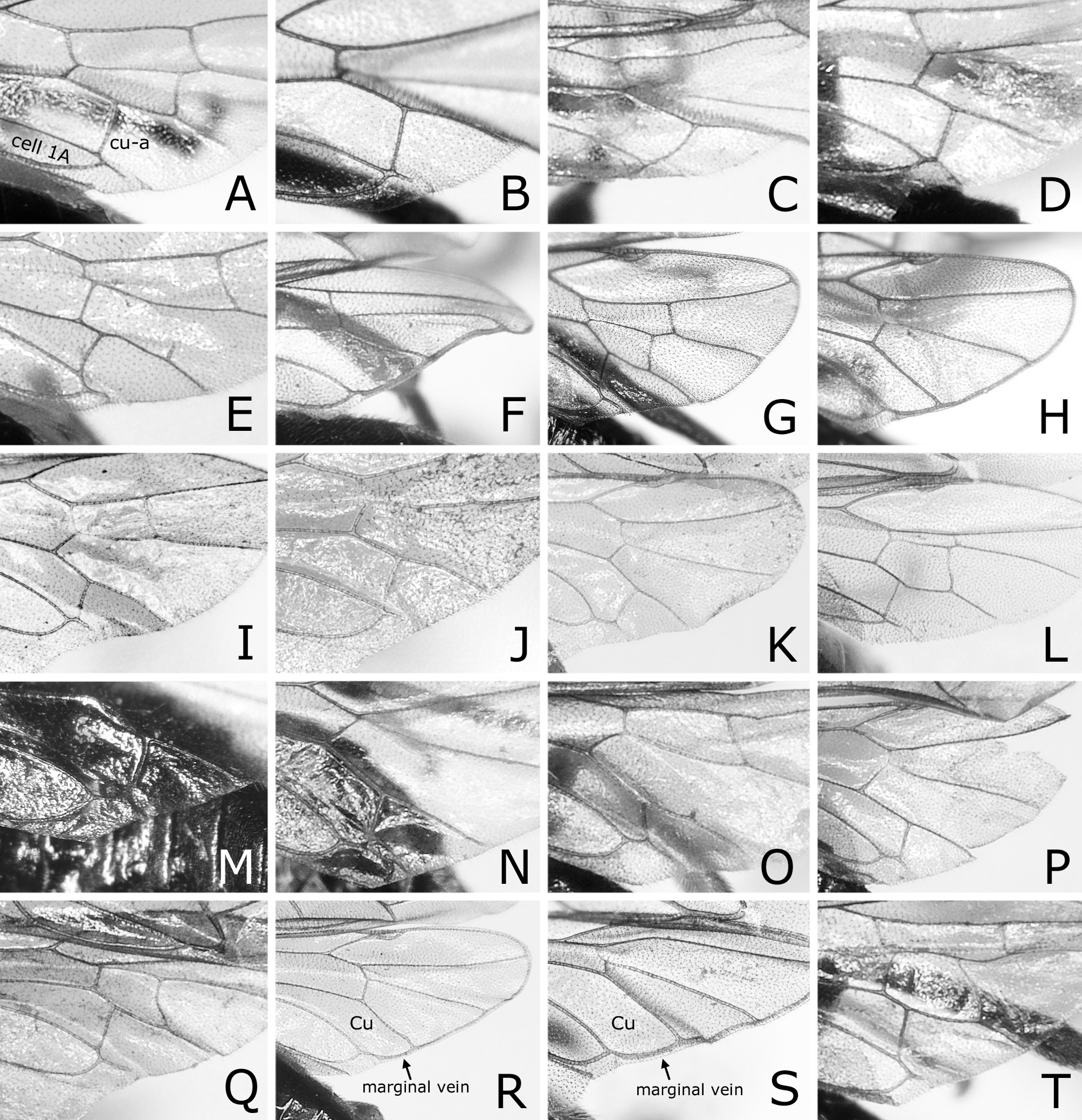
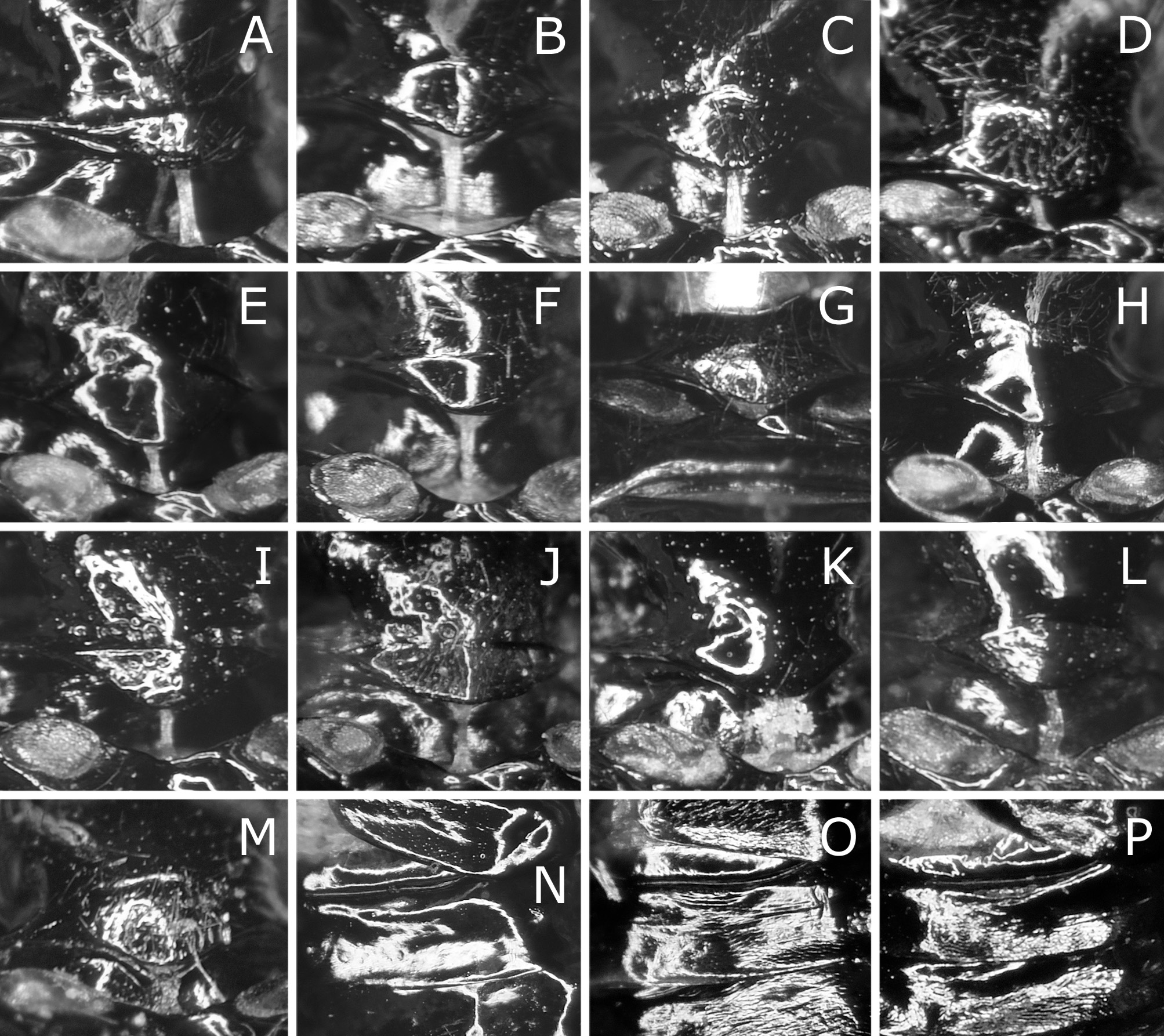
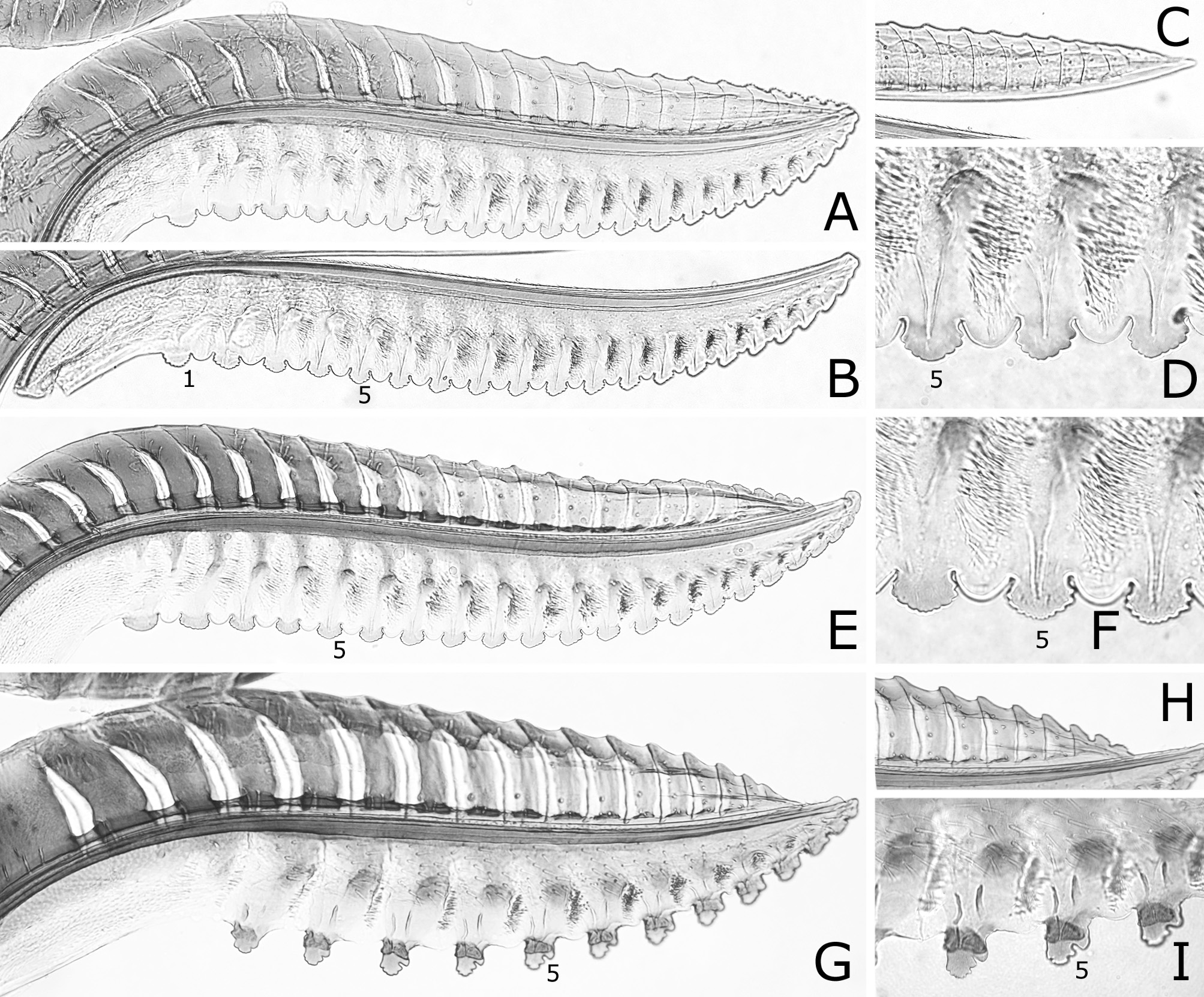
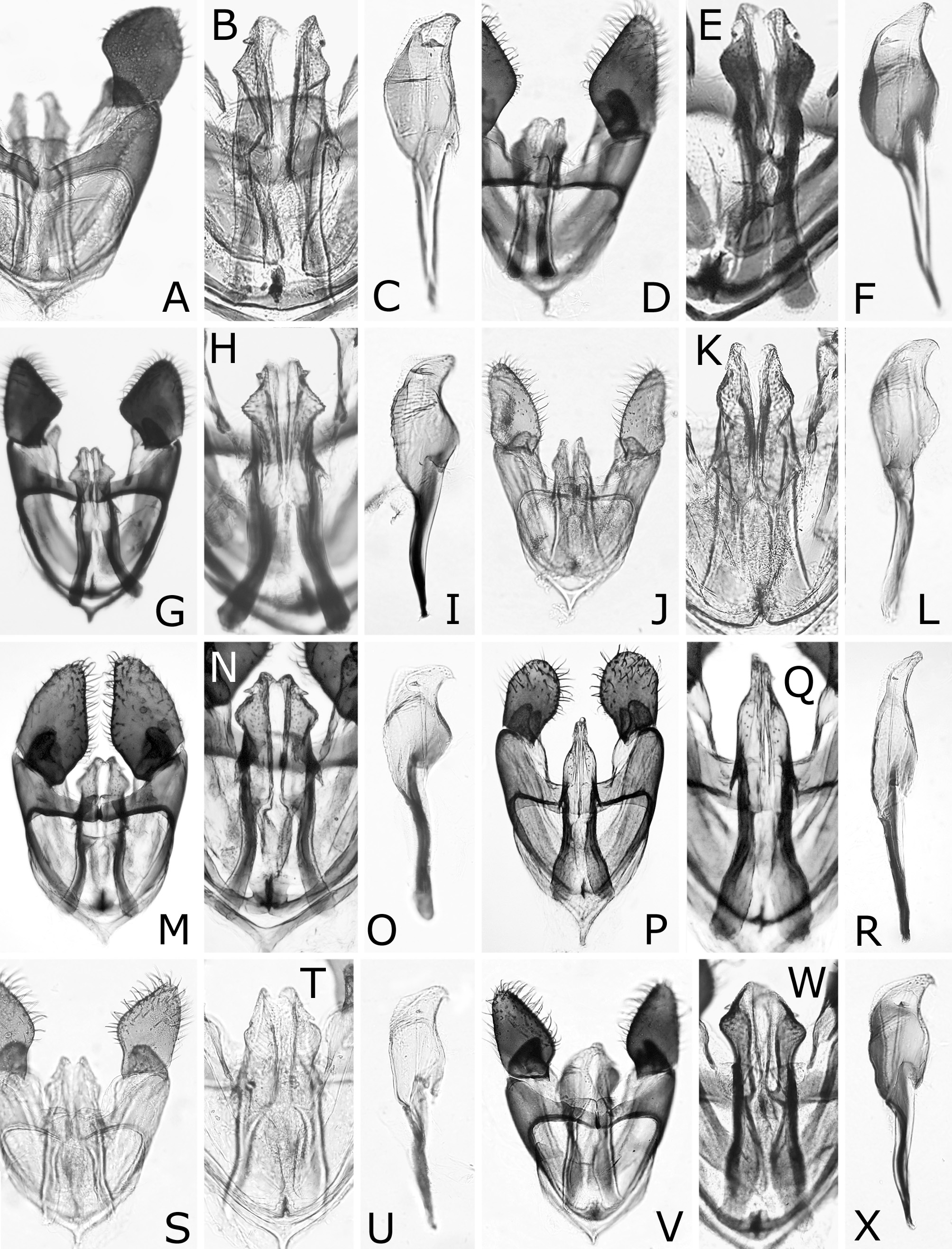
( Figs 1 View FIGURE 1 E–H, M, 3F, P, 4E, F, 5C, 7A–F, 11D–F)
Eriocampoides oishii Takeuchi, 1933: 30 ; Takeuchi, 1936: 160.
Caliroa oishii: Takeuchi, 1949: 48 ; Oishi, 1961: 34; Togashi, 1961: 37; Kim, 1963: 41; Okutani, 1965: 29; Okutani, 1967: 95; Abe & Togashi, 1989: 547; Zhelochovtsev & Zinovjev, 1996: 362; Wei & Nie, 1997: 82; Togashi, 1998: 43; Togashi, 1999: 179; Naito et al., 2004: 28; Wei et al., 2006: 522; Yoshida, 2006: 56; Lelej & Taeger, 2007: 954; Taeger et al., 2010: 366; Lelej, 2012: 78; Lee et al, 2019: 36.
Caliroa View in CoL ? annulipes [sic]: Togashi, 1960: 9. Not Klug (1816).
Caliroa annulipes: Okutani, 1965: 29 , 33. Not Klug (1816).
Caliroa quercivora Togashi, 1999: 177 ; Taeger et al., 2010: 367. Syn. nov.
Redescription: female and male. Length 4.0–5.0 mm in female, 3.5–4.5 mm in male. Black, shiny with colorless reflection ( Fig. 1 View FIGURE 1 E–H). Labrum dark brown to black. Mandible black, apically reddish brown. Palpi black, apically dark brown. Legs black; fore and middle legs white to yellow from apices of femora to tarsi; middle tibia and fore and middle tarsi often apically darkened narrowly; hind leg of female with apex of femur narrowly yellow to brown, tibia white to yellow at least on basal fourth, sometimes almost entirely, and tarsus white to yellow except for apex, rarely only on narrow base; hind leg of male with femur apically yellow to yellow brown widely, tibia yellow to yellow brown, slightly darkened dorsally and apically, and hind tarsus entirely yellow brown to dark brown; tibial spurs yellow, hind tibial spurs often brown; claws brown. Wings uniformly colorless transparent; veins and stigma dark brown to black.
Postocellar area 1.6–1.9 × as wide as length behind lateral ocellus, without anterior groove. Clypeus roundly or angularly emarginated on ventral margin ( Fig. 3F View FIGURE 3 ); depth of emargination 0.2–0.4 × median length of clypeus. Malar space linear, narrower or slightly wider than facet of eye, glabrous. First flagellomere 0.8–0.9 × as long as second and third flagellomeres combined ( Fig. 3P View FIGURE 3 ); apical four flagellomeres combined 1.2–1.3 × as long as first flagellomere. Forewing with joint of vein Rs and crossvein 2r-rs located at apical 0.33–0.44 of anterior margin of cell 1Rs2 ( Fig. 1E, G View FIGURE 1 ); basal corner of cell 1M slightly acute, rarely right-angled. Hind wing of female with joint of vein 1A and crossvein cu-a located basal to apex of cell 1A ( Fig. 4E View FIGURE 4 ); vein 1A extending beyond apex of cell 1A apically; crossvein 2r-m present; crossvein m-cu present or absent. Hind wing of male with marginal vein ( Fig. 4F View FIGURE 4 ); apex of cell 1A very close to wing margin; crossveins 2r-m, m-cu and cu-a absent.
Punctures mostly minute or inconspicuous. Head and thorax mostly smooth. Mesoscutellum posterolaterally with some relatively large punctures along posterior margin ( Fig. 5C View FIGURE 5 ). Mesoscutellar appendage setose, laterally glabrous. Dorsum of abdomen smooth, sometimes partly reticulately microsculptured.
Lance ( Fig. 7A, C, E View FIGURE 7 ) with dorsal margin serrate on apical half; serrations rounded. Lancet ( Fig. 7A, B View FIGURE 7 , D–F) with 19–21 serrulae; ctenidia dark, ventrally pale, and ventrally extending near level of base of serrula; middle serrulae nearly wide oval in outline, with three to five anterior and four to six posterior rounded teeth; areas between middle serrulae distinctly convex and narrower than or about as wide as adjacent serrula.
Male genitalia ( Fig. 11 View FIGURE 11 D–F) in ventral view with harpe widest at basal two fifths, straight or slightly rounded on lateral margin and distinctly convex on medial margin. Penisvalve apically hooked.
Material examined. Type material examined. Holotype or paratopotype of Eriocampoides oishii Takeuchi, 1933 ( Figs 1E, F View FIGURE 1 , 3F, P View FIGURE 3 , 4E View FIGURE 4 , 7 View FIGURE 7 A–D): 1♀, “[Hobara-machi Fukushima-ken [in Japanese)] Takeuchi” [on upper side] “[Oishi (in Japanese)” [on underside], “ HOLOTYPE, Eriocampoides oishii Takeuchi, 1933 , det. A. Shinohara, 1979”. Paratypes of E. oishii : 1♂, “[Hobara-machi, Fukushima-ken (in Japanese)] Takeuchi” [on upper side] “[Oi- shi (in Japanese)]” [on underside], “ ALLOTYPE, Eriocampoides oishii Takeuchi, 1933 , det. A. Shinohara, 1979”; 1♂, “ 29. V. 1917, Minomo [= Minoo, Osaka Pref.], Takeuchi”, “ PARATYPE, Eriocampoides oishii Takeuchi, 1933 , det. A. Shinohara, 1979”.
Takeuchi (1933) described E. oishii based on three females and two males, “ Holotype, ♀, allotopotype, ♂, 1 paratopotype, ♀, Hohara [= Hobara], Fukushimaken, date unknown ( T. ÔISHI); 2 paratypes, ♀ ♂, Minoo near Osaka, May 29, 1917 (K. TAKEUCHI)”. There are no specimens with the type label of Eriocampoides oishii by the nomenclator in Takeuchi’s collection now kept in the National Museum of Nature and Science, Tsukuba. However, we have found the above one female and two males agreeing with the original description in the collection. They are safely considered a part of the type series. The female has the holotype label by A. Shinohara in 1979 (unpublished determination), and it was probably regarded by Togashi (1999) as the holotype. However, there is no reason to consider it the holotype, not the paratopotype. We treat it as indeterminate whether it is the holotype or paratopotype.
Holotype of Caliroa quercivora Togashi, 1999 ( Figs 1G, H View FIGURE 1 , 7E, F View FIGURE 7 ): ♀, “1300~ 1500 m, Mt. Hakusan , Ishikawa Pref., 22. VIII. 1998, I. Togashi ”, “Host: [Mizunara (in Japanese; = Quercus crispula )]”, “ Holotype, Caliroa quercivora sp. nov. ”, “ NSMT-HYM 62270 ” . Paratype of C. quercivora : 1♀, kept beside the holotype, bearing only a small red rectangular paper with nothing written on it.
We regard the female only with a red paper as a paratype of C. quercivora , because Togashi (1999) wrote that “ Holotype and one paratype are deposited in the collection of the National Science Museum (Nat. Hist.), Tokyo [= National Museum of Nature and Science , Tsukuba ]” .
Other material examined. JAPAN—HOKKAIDO: 1♀, Tokachi, Shintoku , on Quercus crispula , 13. VIII. 1993, H. Hara ; 1♀, same data but coll. larva on Q. crispula 13. VIII. 1993, mat. 29–31. VIII., em. 25. V. 1994 ; 2♀ 1♂, same data but coll. larvae 19. VIII. 2011, mat. 22–23. VIII., em. 19–26. IV. 2012, H. Hara ; 2♀ 1♂, Tokachi, Shintoku , Mt. Shintoku-yama, coll. 3 larvae on Q. crispula 19. VIII. 2011, mat. 22. VIII., em. 2–5. IX. 2011, H. Hara ; 1♀ 5♂, Fukagawa, Takadomari, 6–14. VI, 14–24. VI, or 1–16. VII, 2007, Malaise trap, H. Hara ; 2♀, Mikasa, Kayano , 11. VI. 2016, H. Hara ; 1♀, Jozankei , 20. VI. 1932, H. Sugiura ; 1♀, Kutchan , 26. VI. 1932, H. Sugiura ; 1♀, Hakodate, Kikyo, coll. larva on Q. crispula 17. VII. 2008, mat. 19. VII., em. 12. V. 2009, H. Hara. —HONSHU: Yamagata
Pref.: 1♀, Sodeura, 29. VI. 1951, Takeuchi, K, Shirahata.— Niigata Pref.: 1♀, Sado Is., Mt. Kinpoku-san, 7–8. VII. 1936, K. Baba, Y. Suzuki & Z. Sawano [cited by Takeuchi (1936)].— Nagano Pref.: 1♂, Mt. Jonen-dake, 16. VII. 1929, Takeuchi.— Toyama Pref.: 1♂, Kurobe, 21. VI. 1931, Takeuchi.— Ishikawa Pref.: 1♀, Mt. Haku-san, Mt. Rokuman-yama, 22. VII. 1952, Takeuchi, I. Togashi; 1♀, same locality, 21. VII. 1962; 1♀, Mt. Shiritaka-yama, 2. VII. 1954, “ Caliroa ? annulipes Klug. ” [cited by Togashi (1960) as “ Caliroa ? annulipes Klug. ”; material of “ C. annulipes ” of Okutani (1965) (see Hara, 2011)].— Kyoto Pref.: 1♀, Kyoto, Sakyo-ku, Ohara, 16. VI. 1984, T. Mat- sumoto.— Hyogo Pref.: 1♀, Sasayama, Mt. Sakazukigatake, 7. VII. 1951, K. Iwata; 1♂, Sasayama, Okano, 21. VI. 1954, T. Okutani [probably material of Okutani (1965)]; 1♀, Mt. Sengamine, 8. VII. 1962 [probably material of Okutani (1965)].— Tottori Pref.: 1♀, Mt. Daisen, 7. VI. 1933, Takeuchi.
Distribution. Japan: Hokkaido ( Togashi, 1998), Honshu ( Takeuchi, 1933), Sado Is. ( Takeuchi, 1936). Sakhalin ( Zhelochovtsev & Zinovjev, 1996), Russian Far East ( Zhelochovtsev & Zinovjev, 1996); Korea ( Kim, 1963); China, Hubei ( Wei & Nie, 1997).
Bionomics. Host plants: Fagaceae : Quercus acutissima Carruth. ( Oishi, 1961 as “[Kunugi (in Japanese)]”), Q. aliena Blume ( Lee et al, 2019), Q. serrata Murray ( Takeuchi, 1933 as “ Quercus serrata and Quercus glandulifera ”), Q. crispula Blume ( Oishi, 1961 as “[Midunara (in Japanese)]”, Q. variabilis Blume ( Lee et al, 2019).
According to Oishi (1961), this species has two or more generations a year; a female lays an egg below the epidermis of the underside of a leaf from the upper side of a leaf; a larva inhabits the underside of a leaf. He also wrote that up to 40– 50 eggs were laid on a leaf, but we have observed at most three larvae on one leaf. In our observations, larvae were solitary; two or three larvae were very often found on one leaf, but they were separated from each other; mature larvae entered into the soil and made brittle cocoons; hibernation was done in a cocoon.
Remarks. Togashi (1999) described C. quercivora as a species closely resembling C. oishii and separated them by the ratio of length to width of a postocellar area, the shape of an ovipositor sheath and the coloration of a hind tarsus. He wrote that a postocellar area has “length to width ratio of 1.0:1.8” for C. quercivora , but about “1.0:1.4” for C. oishii . However, actually the ratio of the holotype or paratopotype of C. oishii is 1.0:1.9 and very close to the ratio 1.0:1.8 of the holotype of C. quercivora . The shape of an ovipositor sheath and the color of a hind tarsus are not very different between these two species: compare fig. 11 and fig. 13 in Togashi, 1999 for their ovipositor sheaths, and Fig. 1F View FIGURE 1 and Fig. 1H View FIGURE 1 for the color of their hind tarsi. Furthermore, the ovipositor of the holotype or paratopotype of C. oishii and that of the holotype of C. quercivora are almost identical (compare Fig. 7 View FIGURE 7 A–D and Fig. 7E, F View FIGURE 7 ). We consider C. quercivora a junior synonym of C. oishii .
In eastern Palearctic and Oriental species, C. oishii is similar to C. bilobatina Wei, 2002 described from central China in having a black body with colorless reflection, a pale hind tibia and tarsus with dark apices, uniformly colorless transparent wings, a linear malar space and a female hind wing with the joint of vein 1A and crossvein cu-a located basal to the apex of cell 1A. However, they differ as follows: first flagellomere 0.8–0.9 × as long as second and third flagellomeres combined in C. oishii , 0.6 × in C. bilobatina ; serrulae of a lancet wide oval in C. oishii ( Fig. 7D, H View FIGURE 7 ), bilobate in C. bilobatina (fig. 3b, c in Wei, 2002).
In the key to western Palearctic species by Lacourt (2002), C. oishii goes to the couplet 5 or 6, but does not agree with either line in both couplets.
In the key to Nearctic species by Smith (1971), the female of C. oishii goes to the couplet 14 containing the females of C. lobata MacGillivray, 1909 and C. fasciata (Norton, 1864) , but it differs from the latter two species in having uniformly colorless transparent wings and apical four flagellomeres combined 1.2–1.3 × as long as a first flagellomere [wings uniformly, moderately infuscated, sometimes lighter on the apical third, and apical four flagellomeres subequal in length to a first flagellomere in C. lobata ; wings darkened on the basal two thirds and hyaline on the apical third, and apical four flagellomeres together slightly shorter than a first flagellomere in C. fasciata ]. Their lancets also are different (compare Fig. 7 View FIGURE 7 A–F with figs 73 and 75 in Smith, 1971). In Smith’s key, the male of C. oishii will go to the male of C. nyssae Smith, 1971 , but apical four flagellomeres combined is 1.2–1.3 × as long as a first flagellomere in C. oishii , subequal in C. nyssae .

| V |
Royal British Columbia Museum - Herbarium |
| T |
Tavera, Department of Geology and Geophysics |
| VI |
Mykotektet, National Veterinary Institute |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Caliroa oishii ( Takeuchi, 1933 )
| Hara, Hideho & Ibuki, Shinichi 2020 |
Caliroa quercivora
| Taeger, A. & Blank, S. M. & Liston, A. D. 2010: 367 |
| Togashi, I. 1999: 177 |
Caliroa annulipes:
| Okutani, T. 1965: 29 |
Caliroa
| Togashi, I. 1960: 9 |
Caliroa oishii: Takeuchi, 1949: 48
| Lee, J. - W. & Choi, J. - K. & Park, B. 2019: 36 |
| Lelej, A. S. 2012: 78 |
| Taeger, A. & Blank, S. M. & Liston, A. D. 2010: 366 |
| Lelej, A. S. & Taeger, A. 2007: 954 |
| Wei, M. & Nie, H. & Taeger, A. 2006: 522 |
| Yoshida, H. 2006: 56 |
| Naito, T. & Yoshida, H. & Nakamine, H. & Morita, T. & Ikeda, T. & Suzuki, H. & Nakanishi, A. 2004: 28 |
| Togashi, I. 1999: 179 |
| Togashi, I. 1998: 43 |
| Wei, M. & Nie, H. 1997: 82 |
| Zhelochovtsev, A. N. & Zinovjev, A. G. 1996: 362 |
| Abe, M. & Togashi, I. 1989: 547 |
| Okutani, T. 1967: 95 |
| Okutani, T. 1965: 29 |
| Kim, C. W. 1963: 41 |
| Oishi, T. 1961: 34 |
| Togashi, I. 1961: 37 |
| Takeuchi, K. 1949: 48 |
Eriocampoides oishii
| Takeuchi, K. 1936: 160 |
| Takeuchi, K. 1933: 30 |